1. Cilium
2. Flagelle
3. Structure
4. Mouvement
5. Formation
Les microtubules sont des composants du cytosquelette ayant des fonctions importantes dans la physiologie cellulaire. L’échafaudage microtubulaire du cytoplasme est très plastique grâce à la capacité de polymérisation et de dépolymérisation des microtubules. Cependant, tous les microtubules cellulaires ne sont pas soumis à cette phase de contraction ou de croissance. Les cils, les flagelles et les centrioles/corps basaux sont des structures cellulaires contenant des microtubules très stables (nombre et longueur) et très organisés. Dans cette page, nous traitons des cils et des flagelles.
1. Cilium
Les cils sont des protubérances cellulaires fines et longues d’environ 0,25 µm de diamètre et d’environ 10 à 15 µm de longueur, que l’on trouve dans les cellules animales et certaines espèces eucaryotes unicellulaires. Ils sont généralement serrés à la surface libre des cellules épithéliales (figures 1 et 2), comme l’épithélium des voies respiratoires, l’épithélium des canaux reproducteurs, les branchies des poissons et des bivalves, etc. Les cils sont des structures mobiles et leur principale fonction est de déplacer le liquide environnant, comme le mucus de la surface des voies respiratoires, l’eau autour de l’épithélium des branchies, mais aussi l’ovocyte dans le canal de Fallope de la femelle. De nombreux organismes unicellulaires peuvent se déplacer grâce aux cils, et d’autres peuvent les utiliser pour générer des tourbillons d’eau afin d’attraper de la nourriture. Les cils nodaux de l’embryon ont été impliqués dans l’initiation de l’axe gauche-droite au cours du développement embryonnaire des vertébrés. Le mouvement des cils s’apparente à un battement, qui impulse le liquide parallèlement à la surface de la cellule.
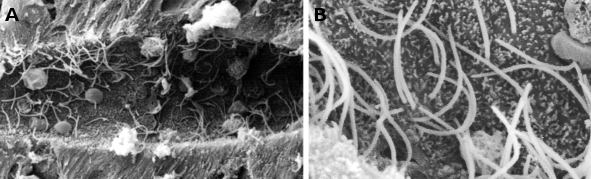
Figure 1. Images de microscopie électronique à balayage montrant le canal central d’une moelle épinière de lamproie. On peut observer de nombreux cils (à plus fort grossissement en B) et de petites microvillosités au niveau du domaine apical des cellules épendimales.
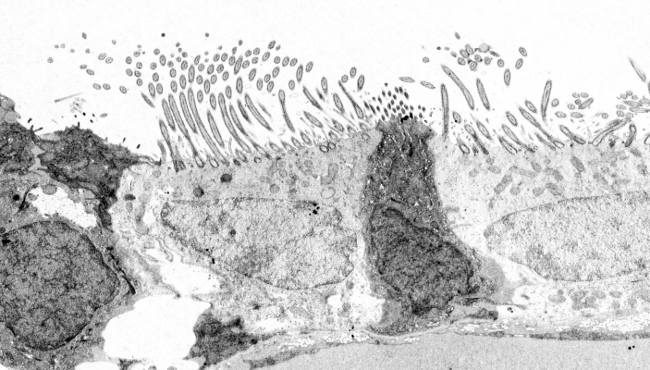
Figure 2. Images de microscopie électronique à transmission de l’épithélium respiratoire. Les cellules avec un cytoplasme clair montrent de nombreux cils dans leur surface apicale.
Il existe des cils qui ne peuvent pas se déplacer, et qui ne sont donc pas destinés au mouvement des liquides. Ces cils sont connus sous le nom de cils primaires. La plupart des cellules étudiées jusqu’à présent (à l’exception des globules rouges) portent des cils primaires : cellules de l’oviducte, neurones, chondrocytes, cellules de l’ectoderme, cellules mésenchymateuses, cellules épithéliales urinaires, hépatocytes, et même cellules en culture. Initialement, les cils primaires étaient considérés comme des cils non fonctionnels. Cependant, de nombreux types de récepteurs et de canaux ioniques se trouvaient dans la membrane ciliaire, de sorte qu’ils étaient considérés comme des structures sensorielles cellulaires. Par exemple, les récepteurs olfactifs se trouvent dans les cils de leurs dendrites, et les segments externes des bâtonnets et des cônes de la rétine sont en fait des cils modifiés. Certains récepteurs sont plus fortement emballés dans la membrane ciliaire que dans d’autres domaines de la membrane plasmique. En outre, il existe une grande variété de molécules à l’intérieur des cils qui jouent un rôle dans la transduction du signal. Le rapport surface/volume plus élevé d’un cilium rend les réponses moléculaires intraciliaires plus intenses et plus efficaces que si elles se trouvaient à l’extérieur du cilium. Outre le signal chimique, les cils primaires peuvent détecter les mouvements de fluide à l’extérieur de la cellule et fonctionner comme des mécanorécepteurs.
2. Les flagelles
Les flagelles sont similaires aux cils, mais ils sont beaucoup plus longs, environ 150 µm, et légèrement plus épais. Ils sont assez moins nombreux que les cils dans les cellules. La fonction principale des flagelles est de déplacer la cellule. Le mouvement du flagelle est différent de celui du cilium car la direction du mouvement est perpendiculaire à la surface de la cellule (et non parallèle), c’est-à-dire la direction de l’axe longitudinal du flagelle. Les flagelles peuvent être fréquemment observés dans les cellules mobiles comme les organismes unicellulaires et les spermatozoïdes.
Structure
Les cils et les flagelles sont des structures complexes contenant plus de 250 protéines différentes. Les deux partagent la même organisation centrale de microtubules et d’autres protéines associées, globalement connues sous le nom d’axonème, et limitées par la membrane plasmique (Figure 3). Outre l’axoneme, il existe de nombreuses molécules solubles à l’intérieur des cils/flagelles qui constituent la matrice. L’axoneme est constitué de 9 paires de microtubules autour d’une autre paire centrale de microtubules. Cette organisation peut s’écrire comme (9 x 2) + 2. Les cils primaires n’ont pas de paire centrale de microtubules. Chaque microtubule de la paire centrale est constitué de 13 protofilaments, mais les microtubules des paires périphériques partagent certains protofilaments entre eux. Ainsi, une paire périphérique est formée par les microtubules A et B. Le microtubule A contient 13 protofilaments et le microtubule B contient 10 ou 11 protofilaments, en partageant 2 ou 3 avec le microtubule A.

Figure 3. Principaux composants moléculaires des cils et des flagelles. Dans les cils primaires, la paire centrale est absente.
L’organisation microtubulaire de l’axonème est le résultat d’un échafaudage de protéines. Douze protéines ont déjà été trouvées comme constituants de l’axonème impliqués dans le maintien de l’organisation des microtubules. Les paires de microtubules périphériques voisines sont reliées entre elles par la nexine. Dans chaque paire, le microtubule A est relié par des rayons protéiques à un anneau central qui contient la paire ou les microtubules centraux. La nexine est une protéine motrice associée aux microtubules périphériques impliqués dans le mouvement des cils et des flagelles.

Figure 4. Ultrastructure d’un cilium d’une cellule épendimale de la moelle épinière. (9+2)x2 signifie 9 paires périphériques et 1 paire centrale de microtubules.
Les microtubules sont polymérisés à partir des corps basaux (figures 3 et 4). Le corps basal est constitué de 9 microtubules triplets formant un cylindre (similaire aux centrioles). Ils sont dépourvus d’une paire centrale de microtubules, ce qui donne (9×3)+0. Dans chaque triplet, seul un microtubule (microtubule A) possède un ensemble complet de protofilaments, tandis que les microtubules B et C en partagent certains entre eux. À partir du corps basal, les microtubules A et B se développent et forment les microtubules périphériques de l’axonème. Juste au-dessus du corps basal, il y a une région des cils connue sous le nom de zone de transition contenant les 9 paires périphériques et aucune paire centrale. Immédiatement après la zone de transition, on trouve la plaque basale, à partir de laquelle la paire centrale de microtubules est polymérisée pour compléter l’axonème. Tous les microtubules ont l’extrémité positive vers l’extrémité du cilium/flagellum. L’extrémité proximale du corps basal (l’interne, ou extrémité négative des microtubules) est ancrée au cytosquelette cellulaire par de longues fibres protéiques appelées radicelles ciliaires
En dehors de l’axonème, les cils/flagelles ont d’autres compartiments. La membrane contient de nombreux récepteurs et canaux pour détecter l’environnement, spécialement dans les cils primaires. La phase fluide de l’intérieur est appelée matrice, qui aide à maintenir l’organisation de l’ensemble de la structure et est responsable de la transduction des informations recueillies par les récepteurs membranaires. D’autres zones distinctes sont le corps basal situé à la base et la partie apicale du cilium/flagellum, qui contient des protéines qui stabilisent les extrémités plus des microbules.
4. Mouvement
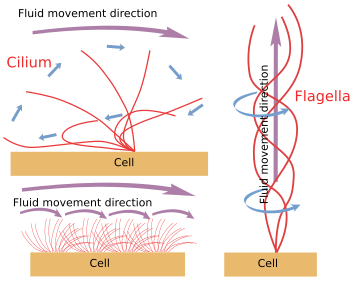
Figure 6. Modèles pour le mouvement des ciliums et des flagelles. Ils génèrent différentes directions de mouvement des fluides.
Lorsque le cilium/flagelle est mécaniquement détaché de la cellule, il continue à se déplacer jusqu’à ce que le stock d’ATP soit épuisé. Cela signifie que le mécanisme de mouvement est intrinsèque (figure 6). En fait, le mouvement est produit par le glissement d’une paire périphérique de microtubules sur le voisin. Les protéines Nexin et spoke empêchent la désorganisation de l’axonème, mais permettent ces mouvements. La dineine est la protéine motrice responsable du mouvement de glissement. L’énergie est fournie par l’ATP. Les dineines sont ancrées avec sa partie globulaire au microtubule A d’une paire périphérique et avec sa partie queue au microtubule B de la paire adjacente. Le mécanisme moléculaire est similaire à celui des dineines cytosoliques, mais au lieu de transporter une cargaison, il déplace un microtubule. Pour un mouvement efficace, une coordination des dineines de l’axoneme est nécessaire. Les ondes calciques à l’intérieur des cils/flagelles peuvent coordonner l’activation des dineines et changer la fréquence du mouvement si nécessaire. Il est à noter que toutes les dineines ne doivent pas être activées en même temps, mais en synchronie.
5. Formation
Lors de la différenciation, les cellules produisent tous les cils et flagelles nécessaires à leur physiologie normale. Cela signifie qu’ils doivent tous être générés à partir de zéro. Les microtubules des axonèmes sont nucléés à partir des microtubules A et B des corps basaux, il faut donc un corps basal par cilium/flagelle. Comment se forment les corps basaux multiples ? Il existe au moins trois façons de produire des corps basaux : a) en utilisant les centrioles comme modèles pour nucléer les corps basaux ; b) à partir d’un matériau amorphe connu sous le nom de deutérosome ; c) chez les plantes, il existe des agrégats protéiques distincts qui peuvent nucléer les corps basaux.
De nombreuses pathologies humaines sont la conséquence de défauts du cilium/flagelle. Elles sont connues sous le nom de ciliopathies, et comprennent la latéralité aléatoire, la mauvaise fermeture du tube neural, la polydactylie, les pathologies kystiques du rein, du foie et du pancréas, la dégénérescence de la rétine, l’obésité et les défauts cognitifs.
Bibliographie
Marshall WF, Nonaka S. 2006. Cilia : tuning in to the cell’s antenna. Current biology. 16:R604-R614.
Satir P, Christensen ST. 2007. Aperçu de la structure et de la fonction des cils des mammifères. Revue annuelle de phisiologie. 69:377-400.