Systématique : Initialement décrit comme Coluber obsoletus par Thomas Say en 1823 à partir d’un spécimen collecté à « Isle au Vache à Council Bluffs sur le fleuve Missouri. » Schmidt (1953) a restreint la localité type à Council Bluffs, Iowa. Dunn (1915a) a été le premier à utiliser le genre Elaphe pour cette espèce mais a épelé l’espèce obsoletus. En 1836, Holbrook a décrit Coluber alleghaniensis à partir d’un spécimen trouvé « au sommet de la Blue Ridge en Virginie ». Ce spécimen et d’autres provenant de New York et des montagnes de la Caroline étaient les premières couleuvres obscures de l’Est qu’il voyait et qui avaient des écailles faiblement carénées. Le nom a été plus tard considéré comme un synonyme junior de E. obsoletus (= obsoleta) par Cope (1900). Callopeltis obsoletus a été utilisé pour cette espèce par Hay (1902), suivant Lonnberg (1894). Sur la base de la congruence des données morphologiques (Burbrink, 2001, Herpetol. Monogr. 15 : 1-53) et mitochondriales (Burbrink et al., 2000, Evolution 54 : 2107-2118), Burbrink a divisé P. obsoletus en trois espèces (P. alleghaniensis, P. obsoletus et P. spiloides) sans aucune sous-espèce. P. alleghaniensis est la seule espèce présente en Virginie.
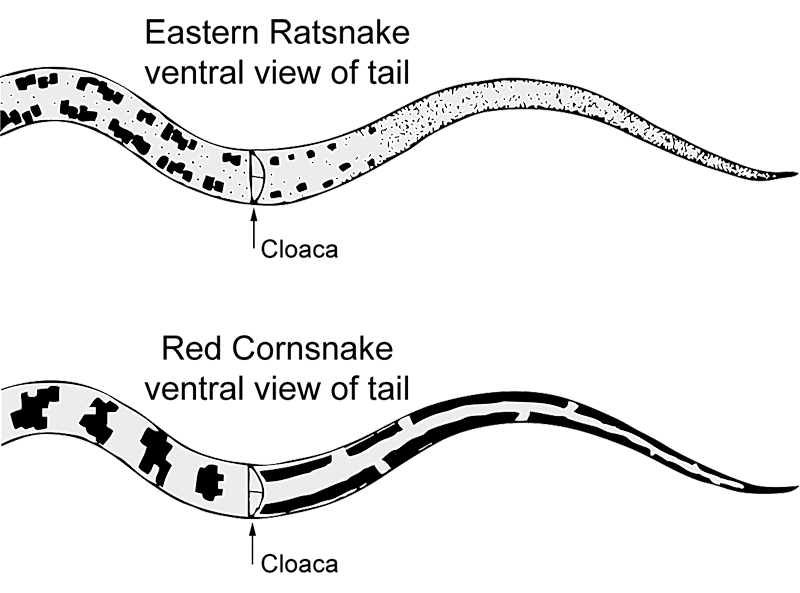
Description : Un grand serpent corpulent atteignant une longueur totale maximale connue de 2 565 mm (101,0 pouces) (Conant et Collins, 1991). En Virginie, la longueur maximale connue du museau-vent (SVL) est de 1 710 mm (67,3 pouces) et la longueur totale maximale est de 2 028 mm (79,8 pouces). Dans cette étude, la longueur de la queue/longueur totale était en moyenne de 16,9 ± 1,6 % (11,2-23,6, n = 209).
Scutellation : Ventrales 214-272 (moy. = 231,8 ± 5,2, n = 232) ; subcaudales 46-94 (moy. = 80,0 ± 7,4, n = 203) ; ventrales + subcaudales 275-348 (moy. = 312,1 ± 9.3, n = 201) ; écailles dorsales lisses latéralement et faiblement carénées médialement, rangées d’écailles généralement 24-27 (77,1%, n = 251) au milieu du corps, mais pouvant être 21-23 ou 28 (22,9%) ; plaque anale non divisée (10,8%) ou divisée ou partiellement divisée (89.2%, n = 232) ; infralabiales 11/11 (47.3%, n = 167), 10/10 ou 10/11 (21.6%), 11/12 ou 12/12 (21.6%), ou autres combinaisons de 9-13 (9.5%) ; supralabiales 8/8 (89.3%, n = 224) ou autres combinaisons de 6-9 (10.7 %) ; loréal présent ; préoculaires 1/1 ; postoculaires 2/2 ; temporaux généralement 2+3/2+3 (52,4 %, n = 229), 2+2/2+3 (13,1 %), 2+2/2+2 (10,5 %), 2+4/ 2+3 (10,0 %), ou autres combinaisons de 1-3/2-6 (14,/4 %).
Coloration et motif : Corps uniformément noir dorsalement chez les adultes ; certains individus présentent de faibles rayures noires sur un corps gris-noir ou un motif juvénile incomplètement masqué (voir ci-dessous) ; venter avec un motif irrégulier en damier noir et blanc entrecoupé de taches noires, le rose remplaçant le blanc chez certains individus ; le motif ventral s’estompe vers l’arrière pour devenir entièrement gris chez les adultes plus âgés, mais un motif poivré noir et blanc chez les individus plus jeunes ; menton et partie antérieure du venter du cou blanc uni ; un pigment blanc est également présent sur la moitié inférieure des supralabiales. Le corps en coupe transversale imaginaire a la forme d’une miche de pain, avec un venter plat.
Dimorphisme sexuel : Il n’y a pas de différences sexuelles dans la couleur ou le motif. La SVL des mâles adultes (908-1 710, moy. = 1 182,1 ± 187,1, n = 124) était similaire à celle des femelles adultes (910-1 590, moy. = 1 136,1 ± 156,0, n = 54). L’indice de dimorphisme sexuel était de -0,04. Les mâles ont atteint des longueurs totales plus longues (jusqu’à 2 028 mm) que les femelles (jusqu’à 1 884 mm). La longueur de la queue par rapport à la longueur totale était plus élevée chez les mâles (moy. = 17,5 ± 1,4 %, 15,1-23,6, n = 107) que chez les femelles (moy. = 15,7 ± 1,3 %, 11,2-18,0, n = 47). Le nombre moyen d’écailles ventrales était légèrement plus élevé chez les femelles (233,4 ± 7,3, 214-272, n = 50) que chez les mâles (230,7 ± 3,9, 220-246, n = 112). Le nombre moyen d’écailles subcaudales était plus élevé chez les mâles (82,5 ± 5,7, 64-94, n = 96) que chez les femelles (74,3 ± 8,6, 46-88, n = 45), mais le nombre moyen de ventrales + subcaudales était similaire entre les sexes (mâles 313,3 ± 7,6, 290-337, n = 95 ; femelles 308,4 ± 11,5, 275-348, n = 44).
Juvéniles : Contrairement aux adultes, les juvéniles présentent un fort motif de taches noires à marron foncé dorsalement (moy. = 33,5 ± 2,9, 28-40, n = 36) sur un corps poivré noir et blanc à gris. Les taches antérieures ont des projections antérieures et postérieures sur les coins. Le venter brun et blanc forme un damier. Il y a une bande brune distincte qui commence devant l’œil et va jusqu’au bord de la bouche. Ces bandes sont reliées par une bande brune sur le dos de la tête. Le ventre de la queue présente une bande sombre irrégulière de chaque côté. Le motif juvénile est généralement obscurci à partir d’une longueur hors tout d’environ 650 mm. Certains jeunes adultes peuvent présenter un léger motif de tache dorsale, qui peut également être observé chez les adultes adultes dans certaines régions (voir « Variation géographique »). À l’éclosion, les juvéniles de Virginie avaient en moyenne 284,6 ± 14,2 mm de LCS (258-318, n = 48) et 343,9 ± 17,3 mm de longueur totale (317-391, n = 47), et pesaient de 9,4 à 13,2 g (moyenne = 11,7 ± 1,2, n = 21).
Espèces pouvant prêter à confusion : Les couleuvres obscures de l’Est adultes peuvent être confondues avec les Coluber constrictor adultes ; cependant, ces derniers ont des écailles entièrement lisses et un corps rond en coupe transversale, et le pigment blanc est confiné au menton. Les C. constrictor juvéniles sont dépourvus de la bande œil-mâchoire, du motif en damier sur le venter, des projections sur les taches dorsales antérieures et des bandes sur le venter de la queue. Ils possèdent également 1,5 à 2 fois le nombre de taches dorsales. Les juvéniles sont parfois confondus avec les petits Agkistrodon contortrix, mais les Copperheads de l’Est possèdent des bandes transversales brunes en forme de sablier et un bout de queue jaune.
Variation géographique : Les Pantherophis alleghaniensis adultes sont uniformément noirs dorsalement dans la majeure partie de la Virginie. Les individus de l’extrême sud-est et des environs du comté de Greensville présentent des traces variables des quatre bandes noires longitudinales sur un fond gris foncé. Certains individus du sud-ouest de la Virginie, notamment dans les comtés de Pulaski et de Washington, conservent une partie du motif juvénile, mais la variation est grande. Le nombre moyen d’écailles ventrales ne varie pas de façon significative entre les régions physiographiques, allant de 229,3 ± 4,8 (220-237, n = 12) sur la côte Est à 233,9 ± 3,4 (230-243, n = 13) dans le Piedmont inférieur. Le nombre moyen d’écailles subcaudales était le plus faible dans les populations situées au sud-ouest de la New River dans la région de Ridge and Valley (74,9 ± 9,6, 54-82, n = 14) et le plus élevé (82,8 ± 6,1, 60-92, n = 35) dans le nord du Piémont. Le nombre moyen de ventrales + subcaudales suivait un schéma similaire (SW Ridge and Valley 306,8 ± 11,3, 286-318, n = 14 ; N Piedmont 315,6 ± 6,9, 296-328, n = 35).
Biologie : Les couleuvres obscures orientales sont les serpents les plus couramment observés en Virginie. Elles sont terrestres et arboricoles. Elles sont présentes dans de nombreux types d’habitats, notamment les zones agricoles, la plupart des types de forêts de feuillus, les boisés urbains isolés et les zones humides boisées. On trouve souvent ces serpents dans les granges et les vieux bâtiments où leurs proies principales, les petits rongeurs, se trouvent en abondance. Hutchison (1956) en a trouvé une dans la bouche d’une grotte. Les couleuvres obscures de l’Est sont diurnes et nocturnes. Elles sont souvent actives juste après le coucher du soleil. Pendant cette période, elles se déplacent sur des distances considérables et beaucoup sont tuées par des véhicules sur les routes. Cette espèce subit de grandes pertes chaque année à cause de cette source de mortalité. Les données du musée concernant le P. alleghaniensis de Virginie indiquent une période d’activité allant du 3 avril au 8 décembre. On sait que l’activité est brève pendant les mois d’hiver, mais elle dépend des conditions météorologiques. Clifford (1976) les a trouvées actives de mai à septembre dans le comté d’Amelia, et Bazuin (1983) a noté une saison d’activité du 11 mars au 8 novembre dans le comté de Louisa. La température corporelle des serpents actifs était de 25,0-30,6°C (moy. = 27,6 ± 1,9, n = 11). Les serpents trouvés sous des objets cachés étaient de 15,0-18,9°C (moy. = 17,5 ± 2,2, n = 3).
Les rongeurs, les oiseaux et les œufs d’oiseaux sont les proies préférées de P. alleghaniensis. Les espèces suivantes ont été enregistrées (Uhler et al, 1939 ; cette étude) : mammifères – tamias striés (Tamias striatus), écureuils gris (Sciurus carolinensis), sylvestres de l’Est (Sylvilagus floridanus), écureuils volants du Sud (Glaucomys volans), le campagnol des prés (Microtus pennsylvanicus), le campagnol des pins (Microtus pinetorum), la souris à pieds blancs (Peromyscus leucopus) et la musaraigne à queue courte (Blarina brevicauda) ; les oiseaux : le merle d’Amérique (Turdus migratorius), le merle bleu de l’Est et ses œufs (Sialia sialis), le pic à ventre jaune (Sphyrapicus varius), le pic mineur (Picoides pubescens), le merle gris (Dumetella carolinensis), la grive brune (Toxostoma rufum), le tarier des prés (Sturnella magna), le bruant chanteur (Melospiza melodia), le colibri à gorge rubis (Archilochus colubris), la grive d’eau (Seiurus noveboracensis), une fauvette non identifiée, un « merle » et des œufs de tétras ou de cailles ; des reptiles : Des scinques à cinq lignes (Plestiodon fasciatus), des serpents non identifiés et des œufs de serpents ; des amphibiens : Lithobates spp. Le serpent qui a mangé le grimpereau brun avait consommé un parent et un oisillon. De nombreux oiseaux capturés étaient des oisillons. Les autres proies enregistrées pour les serpents de Virginie sont les lézards de clôture orientaux (Sceloporus undulatus) (Richmond et Goin, 1938) et les hirondelles de rivage (Riparia riparia) (Blem, 1979). Des types de proies supplémentaires pour cette espèce ont été répertoriés dans Brown (1979) et Ernst et Barbour (1989b). Il existe de nombreuses observations de P. alleghaniensis grimpant aux arbres pour s’attaquer aux oiseaux, à leurs œufs et à leurs oisillons. Ces serpents sont occasionnellement vus en train de manger des œufs de poulets domestiques et parfois des objets qui leur ressemblent. Une fois attrapées, les proies sont tuées par constriction, bien que les œufs soient avalés puis brisés dans la gorge. Une étude menée dans le comté de Warren a montré que les individus de P. alleghaniensis, qui chassent par l’olfaction et la vision, consomment plus de campagnols des prés (Microtus pennsylvanicus) mâles territoriaux et femelles allaitantes que les femelles non allaitantes, en raison du comportement défensif accru de ces groupes (Madison, 1978). Les prédateurs de P. alleghaniensis comprennent les éperviers (Buteo spp.), les grands-ducs d’Amérique (Bubo virginianus) et les chats domestiques en liberté (Mitchell et Beck, 1992 ; C. H. Ernst, comm. pers.).
La couleuvre obscure orientale est ovipare et pond une couvée de 5 à 19 œufs par an (moyenne = 11,3 ± 3,3, n = 24). Les pontes de 21 P. alleghaniensis du nord de la Virginie étaient en moyenne de 19,4 œufs (17-24 ; C. H. Ernst, comm. pers.). Les sites naturels de ponte comprennent des arbres creux, debout ou tombés, des tas de compost et de paillis, des tas de sciure et des bûches en décomposition. Certains sites sont utilisés à plusieurs reprises par P. alleghaniensis (Bader, 1984). Les combats entre mâles précèdent parfois l’accouplement (Mitchell, 1981b). Les dates d’accouplement connues se situent entre le 26 mai et le 9 juin. Les dates de ponte enregistrées sont du 3 juin au 17 juillet (Tuck et al., 1971 ; W. H. Martin, comm. pers. ; C. A. Pague, comm. pers. ; J. C. Mitchell, obs. pers.). Les oeufs mesuraient en moyenne 42,8 ± 6,2 x 24,7 ± 4,3 mm (longueur 34,0-57,3, largeur 19,3-39,5, n = 61) et pesaient 9,0-13,1 g (moy. = 11,6 ± 0,8, n = 26). Tous les mâles et les femelles matures que j’ai mesurés avaient plus de 900 mm de LCS. La durée de l’incubation était de 60 à 65 jours, et les éclosions ont émergé du 30 août au 30 septembre.
C’est le serpent le plus fréquemment rencontré en Virginie. Sur 278 serpents enregistrés sur une période de 4 ans dans le comté d’Amelia, 105 étaient des couleuvres obscures de l’Est (Clifford, 1976). Dans les Blue Ridge Mountains, Martin (1976) a noté que 76 des 545 serpents qu’il a trouvés sur les routes étaient de cette espèce. Shekel et al. (1980) ont trouvé une densité de 0,23 serpent par hectare au Maryland, et Fitch (1963b) en a trouvé 1 par hectare au Kansas. Les couleuvres adultes occupent un domaine vital pouvant atteindre 600 m de diamètre, et les mêmes domaines vitaux sont occupés pendant de nombreuses années, voire toute la vie (Stickel et al., 1980). Les distances maximales de déplacement sont de plus de 1 300 m. Ces serpents utilisent généralement un site de couverture à plusieurs reprises pendant la saison active et le même hibernaculum pendant des années. Les hibernacula les plus courants sont les arbres creux et les souches. Ceux-ci abritent généralement des serpents isolés. On sait que les couleuvres obscures se regroupent en hiver, parfois dans les mêmes tanières où hibernent les copperheads de l’Est (Agkistrodon contortrix). Un tel site a été découvert le 15 avril 1967 dans un tas de bois pourri dans le comté de Loudoun par W. H. Martin (comm. pers.).
Remarques : D’autres noms communs dans la littérature de Virginie sont serpent pilote (Cope, 1900), couleuvre noire d’Alleghany (Hay, 1902), couleuvre noire écailleuse ou rouillée et couleuvre pilote (Dunn, 1915a), couleuvre noire des arbres (Dunn, 1936), couleuvre noire des montagnes (Burch, 1940), couleuvre noire pilote (Burch, 1940 ; Conant, 1945) et serpent poulet (Linzey et Clifford, 1981).
La croyance selon laquelle les couleuvres obscures de l’Est et les cuivres de l’Est s’hybrident et produisent une progéniture venimeuse et ressemblant aux couleuvres obscures de l’Est est un mythe. Ces deux espèces ne sont que faiblement apparentées (familles différentes), ce qui indique que l’accouplement compatible et la production d’une progéniture viable sont hautement improbables. D’autres histoires concernant des serpents noirs (vraisemblablement des couleuvres obscures) sont monnaie courante. Beck (1952) mentionne plusieurs mythes auxquels croyaient les habitants du comté de Rappahannock en 1948, notamment que les couleuvres obscures de l’Est étaient capables de tuer les crotales avec des herbes, de se tenir sur leur queue, de traire les vaches et de charmer les oiseaux et les enfants. Le mythe selon lequel les ratsnakes de l’Est guident les copperheads et les crotales vers la sécurité peut avoir donné lieu au nom de « pilote » pour cette espèce (Ernst et Barbour, 1989b).
Des spécimens albinos ou partiellement albinos de P. alleghaniensis ont été signalés dans trois endroits en Virginie : Westmoreland County (Hensley, 1959), Rockbridge County (Carroll, 1950), et Washington County (D. W. Ogle, comm. pers.). Ces deux derniers serpents ont conservé le motif juvénile mais n’avaient pas de pigment noir.
Mitchell et al. (1982) ont signalé le comportement inhabituel d’autophagie (autoconsommation) chez un P. alleghaniensis juvénile du comté de Chesterfield. Le serpent avait été soumis à un changement soudain de température, a commencé à mordre sa propre queue, l’a avalée complètement (avec autant du reste du corps qu’il le pouvait), et est mort. Il en est résulté trois serpentins concentriques, un à l’extérieur et deux à l’intérieur, d’un diamètre de 4 cm.
Les couleuvres obscures orientales pénètrent parfois dans les transformateurs électriques, s’électrocutent et provoquent des pannes de courant. Un tel incident a laissé 13 000 foyers sans électricité dans le comté de Chesterfield (Richmond Times-Dispatch, 29 mai 1991).
Conservation et gestion : L’abondance et la présence répandue de cette espèce, ainsi que sa capacité à survivre dans une grande variété d’habitats, indiquent que P. alleghaniensis a actuellement besoin de peu de protection et de gestion active. Cependant, des centaines d’individus sont tués sur les autoroutes de Virginie chaque année, une source de mortalité que l’éducation du public sur les serpents pourrait aider à réduire. On ne sait pas comment cette espèce réagit à la fragmentation de l’habitat. La gestion pour la présence continue de cette espèce comprend le maintien de grands peuplements de forêts de feuillus.