1. Cilium
2. Flagellum
3. Felépítés
4. Mozgás
5. Kialakulás
A mikrotubulusok a sejtfiziológiában fontos funkcióval rendelkező citoszkeleton komponensek. A citoplazma mikrotubulus-váza a mikrotubulusok polimerizációs és depolimerizációs képességének köszönhetően rendkívül plasztikus. Azonban nem minden sejtmikrotubulus van ebben a zsugorodó vagy növekvő szakaszban. A csillók, a flagellák és a centriolák/alaptestek nagyon stabil (számuk és hosszuk) és magasan szervezett mikrotubulusokat tartalmazó sejtszerkezetek. Ezen az oldalon a csillókkal és a flagellákkal foglalkozunk.
1. A csillók és a flagellák. Cilium
A csillók vékony és hosszú, kb. 0,25 µm átmérőjű és kb. 10-15 µm hosszúságú sejtnyúlványok, amelyek állati sejtekben és egyes egysejtű eukarióta fajokban találhatók. Általában szorosan elhelyezkednek a hámsejtek szabad felszínén (1. és 2. ábra), például a légutak hámjában, a szaporodási csatornák hámjában, a halak és kagylók kopoltyújában stb. A csillók mozgékony struktúrák, és fő funkciójuk a környező folyadék mozgatása, mint például a légutak felszínének nyálkája, a kopoltyúhám körüli víz, de a női petevezetékben a petesejt is. Számos egysejtű élőlény képes a csillók által hajtott mozgásra, mások pedig a táplálék befogásához szükséges vízörvények létrehozására használják őket. Az embriócsomó csillók a gerincesek embrionális fejlődése során a bal-jobb tengely elindításában játszanak szerepet. A csillók mozgása olyan, mint a lüktetés, amely a folyadékot a sejtfelszínnel párhuzamosan mozgatja.
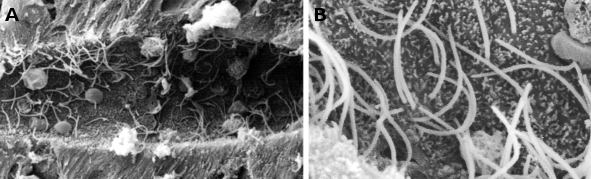
1. ábra. Pásztázó elektronmikroszkópos felvételek, amelyek a lámpás gerincvelő központi csatornáját mutatják. Sok csilló látható (nagyobb nagyításban B-ben) és kis mikrovillák az ependimalis sejtek apikális tartományában.
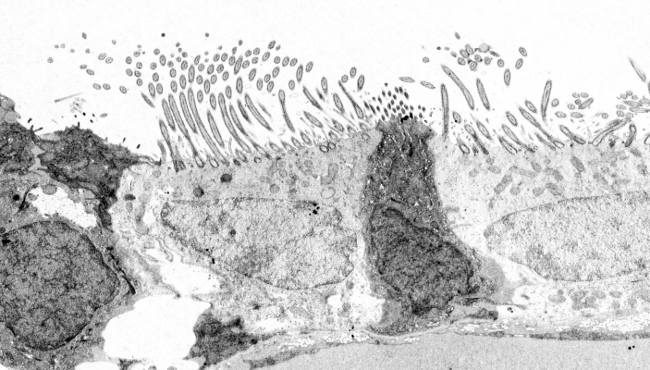
2. ábra. Transzmissziós elektronmikroszkópos felvételek a légzőhámról. A tiszta citoplazmájú sejtek apikális felszínén sok csillószálat láthatunk.
Vannak olyan csillók, amelyek nem tudnak mozogni, ezért nem folyadék mozgatására szolgálnak. Ezeket a csillókat elsődleges csillóknak nevezzük. Az eddig vizsgált sejtek többsége (a vörösvértestek kivételével) primer csillókkal rendelkezik: petefészek sejtek, neuronok, kondrociták, ektoderma sejtek, mesenchymasejtek, húgyhámsejtek, hepatociták és még tenyésztett sejtek is. Kezdetben az elsődleges csillókról úgy gondolták, hogy nem funkcionális csillók. A csillóhártyán azonban számos receptortípust és ioncsatornát találtak, ezért sejtérzékelő struktúráknak tekintették őket. Például a szagló receptorok a dendritek csillóiban találhatók, és a retinák pálcikáinak és kúpjainak külső szegmensei valójában módosított csillók. Egyes receptorok nagyobb sűrűségben helyezkednek el a csillómembránban, mint más plazmamembrán-doménekben. Ezenkívül a csillók belsejében sokféle molekula található, amelyek jelátviteli szerepben vesznek részt. A cilium nagyobb felület/térfogat aránya intenzívebbé és hatékonyabbá teszi a ciliumon belüli molekuláris válaszokat, mintha azok a ciliumon kívül lennének. A kémiai jelek mellett a primer csillók érzékelhetik a sejten kívüli folyadékmozgást, és mechanoreceptorként működhetnek.
2. Az elsődleges csillók a sejten kívüli folyadékmozgást is érzékelhetik. Flagellum
A flagellumok hasonlóak a csillókhoz, de sokkal hosszabbak, körülbelül 150 µm hosszúak és kissé vastagabbak. A sejtekben meglehetősen ritkábban fordulnak elő, mint a csillók. A flagellák fő funkciója a sejt mozgatása. A flagellumok mozgása különbözik a ciliumokétól, mert a mozgás iránya merőleges a sejtfelszínre (nem párhuzamos), vagyis a flagellum hossztengelyének iránya. A flagellák gyakran megfigyelhetők olyan mozgékony sejtekben, mint az egysejtűek és a spermiumok.”
Szerkezet
A csillók és a flagellák összetett szerkezetek, amelyek több mint 250 különböző fehérjét tartalmaznak. Mindkettőnek azonos a központi mikrotubulus szerveződése és más kapcsolódó fehérjék, amelyeket összességében axonémának nevezünk, és a plazmamembrán korlátozza őket (3. ábra). Az axonémen kívül számos oldható molekula található a csillók/lagellák belsejében, amelyek a mátrixot alkotják. Az axonémet 9 pár mikrotubulus alkotja egy másik központi mikrotubuluspár körül. Ezt a szerveződést úgy lehet leírni, hogy (9 x 2) + 2. A primer csillókból hiányzik a központi mikrotubuluspár. A központi pár minden egyes mikrotubulusát 13 protofilamentum alkotja, de a perifériás párok mikrotubulusai néhány protofilamentumot megosztanak egymással. Így a perifériás párt az A és B mikrotubulusok alkotják. Az A mikrotubulus 13 protofilamentumot tartalmaz, a B mikrotubulus pedig 10 vagy 11 protofilamentumot, amelyekből 2 vagy 3 protofilamentumot megoszt az A mikrotubulussal.

3. ábra. A csillók és flagellák fő molekuláris komponensei. A primer csillókban a központi pár hiányzik.
Az axonéma mikrotubulus szerveződése egy fehérjékből álló állványzat eredménye. Az axonéma alkotórészeként már tizenkét fehérjét találtak, amelyek részt vesznek a mikrotubulus szerveződés fenntartásában. A szomszédos perifériás mikrotubuluspárokat nexin köti össze egymással. Minden párban az A mikrotubulus fehérje küllőkkel kapcsolódik egy központi gyűrűhöz, amely a központi párt vagy mikrotubulusokat tartalmazza. A dinein a perifériás mikrotubulusokhoz kapcsolódó motorfehérje, amely részt vesz a csillók és a flagellák mozgásában.

4. ábra. A gerincvelő egyik ependimalis sejtjének ciliumának ultrastruktúrája. A (9+2)x2 9 perifériás és 1 központi mikrotubuluspárt jelent.
A mikrotubulusok bazális testekből polimerizálódnak (3. és 4. ábra). A bazális test 9 triplett mikrotubulusból áll, amelyek egy hengert alkotnak (hasonlóan a centriolákhoz). Hiányzik belőlük egy központi mikrotubuluspár, tehát (9×3)+0. Minden triplettben csak egy mikrotubulus (A mikrotubulus) rendelkezik teljes protofilamentumkészlettel, míg a B és C mikrotubulusok néhányat megosztanak egymás között. A bazális testből az A és B mikrotubulusok nőnek ki, és alkotják az axonéma perifériás mikrotubulusait. Közvetlenül a bazális test felett a csillóknak van egy átmeneti zónának nevezett régiója, amely a 9 perifériás párt tartalmazza, és nincs központi pár. Közvetlenül az átmeneti zóna után található a bazális lemez, amelyből a központi mikrotubuluspár polimerizálódik az axonéma befejezéséhez. Minden mikrotubulusnak a plusz vége a cilium/flagellum csúcsa felé van. A bazális test proximális vége (a belső, vagy a mikrotubulusok mínusz vége) a sejt citoszkeletonjához rögzül hosszú fehérjeszálakkal, az úgynevezett ciliáris gyökerek
Az axonémen kívül a csillóknak/lagelláknak más rekeszei is vannak. A membrán számos receptort és csatornát tartalmaz a környezet érzékelésére, különösen a primer csillókban. A belső tér folyékony fázisát mátrixnak nevezzük, amely segít az egész szerkezet szervezettségének fenntartásában, és a membránreceptorok által gyűjtött információk továbbításáért felelős. További elkülönülő területek a bazális test, amely az alapban található, és a cilium/flagellum apikális része, amely a mikrobolyhok plusz végeit stabilizáló fehérjéket tartalmaz.
4. A ciliumok és az apikális részek (cilium/flagellum) a bazális testet alkotják. Mozgás
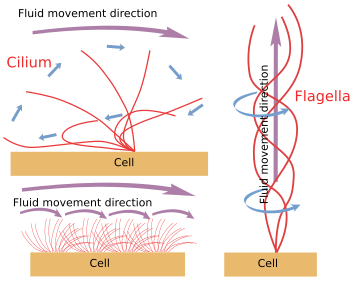
6. ábra. A ciliumok és flagellák mozgásának modelljei. Ezek különböző folyadékmozgási irányokat generálnak.
Amikor a ciliák/lagellák mechanikusan leválnak a sejtről, addig mozognak, amíg az ATP-raktár ki nem merül. Ez azt jelenti, hogy a mozgásmechanizmus intrinzik (6. ábra). Tulajdonképpen a mozgást egy perifériás mikrotubuluspár szomszédon való elcsúszása hozza létre. A Nexin és a spoke fehérjék megakadályozzák az axonéma dezorganizációját, de lehetővé teszik ezeket a mozgásokat. A Dinein a csúszómozgásért felelős motorfehérje. Az energiát az ATP biztosítja. A dineinek globuláris részükkel az egyik perifériás pár A mikrotubulusához, farokrészükkel pedig a szomszédos pár B mikrotubulusához rögzülnek. A molekuláris mechanizmus hasonló a citoszolikus dineinekéhez, de rakomány szállítása helyett egy mikrotubulust mozgat. A hatékony mozgáshoz az axonéma dineinek koordinációjára van szükség. A cilia/flagella belsejében lévő kalciumhullámok koordinálhatják a dineinek aktiválását, és szükség esetén megváltoztathatják a mozgás frekvenciáját. Megjegyzendő, hogy nem minden dineint kell egyszerre, hanem szinkronban aktiválni.
5. A dineinek aktiválódása a szinkronban történik. Kialakulás
A differenciálódás során a sejtek a normális fiziológiájukhoz szükséges összes csillószálat és flagellát létrehozzák. Ez azt jelenti, hogy mindegyiket a semmiből kell létrehozni. Az axonémák mikrotubulusai a bazális testek A és B mikrotubulusaiból nukleálódnak, így ciliánként/lagellánként egy bazális testre van szükség. Hogyan alakulnak ki a többszörös bazális testek? A bazális testek keletkezésének legalább három módja van: a) centriolák felhasználásával, mint sablonok a bazális testek nukleációjához; b) deuteroszóma néven ismert amorf anyagból; c) növényekben vannak külön fehérjeaggregátumok, amelyek képesek bazális testek nukleációjára.
Számos emberi kórkép a cilium/flagellák hibáinak következménye. Ezeket ciliopátiáknak nevezzük, és ide tartozik a véletlenszerű lateralitás, az idegcső helytelen záródása, a polydaktíliák, a cisztás vese-, máj- és hasnyálmirigy-kórképek, a retina degenerációja, az elhízás és a kognitív defektusok.
Bibliográfia
Marshall WF, Nonaka S. 2006. Cilia: a sejt antennájának hangolása. Current biology. 16:R604-R614.
Satir P, Christensen ST. 2007. Az emlősök csillók szerkezetének és működésének áttekintése. Annual review of phisiology. 69:377-400.