1. 繊毛
2. 鞭毛
3. 構造
4. 運動
5. 形成
微小管は細胞骨格の構成要素であり、細胞生理学において重要な機能を持つ。 細胞質の微小管の足場は、微小管の重合・脱重合能により、高度に可塑的である。 しかし、すべての細胞の微小管がこの収縮期や成長期の下にあるわけではありません。 繊毛、鞭毛、遠心体/基底体などは、非常に安定(数と長さ)かつ高度に組織化された微小管を含む細胞構造体である。 このページでは、繊毛と鞭毛を取り上げています。 繊毛
繊毛は直径約0.25μm、長さ約10~15μmの細くて長い細胞の突起で、動物細胞や一部の単細胞真核生物種に見られることがある。 通常、呼吸器上皮、生殖管上皮、魚類や二枚貝のエラなど、上皮細胞の自由表面に密生している(図1、2)。 繊毛は運動性の構造体で、その主な機能は、呼吸器表面の粘液や鰓の上皮の周りの水、さらには雌の卵管内の卵子のように、周囲の液体を移動させることである。 単細胞生物の多くは繊毛を推進力として動くことができるが、餌を捕らえるために水の渦を発生させるために繊毛を利用するものもある。 また、脊椎動物の胎生期における左右軸の形成には、胚節繊毛が関与していることが分かっている。
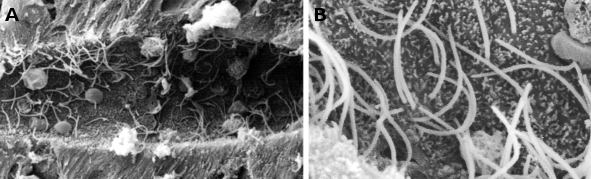
Figure 1. ヤツメウナギの脊髄の中心管を示す走査型電子顕微鏡画像。
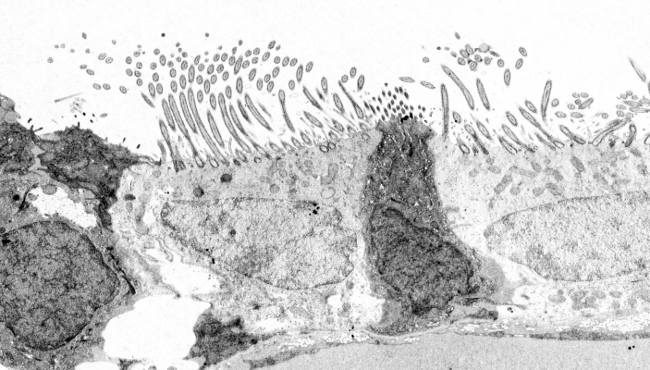
図2. 呼吸器上皮の透過型電子顕微鏡像。 細胞質が透明な細胞は、頂膜表面に多くの繊毛が見られる。
移動できない繊毛もあり、そのため液体運動を目的としていない。 このような繊毛は一次繊毛と呼ばれる。 卵管細胞、神経細胞、軟骨細胞、外胚葉細胞、間葉系細胞、尿路上皮細胞、肝細胞、そして培養細胞など、これまで研究されたほとんどの細胞(赤血球を除く)が一次繊毛を持っている。 当初、一次繊毛は非機能的な繊毛であると考えられていた。 しかし、繊毛膜には多くの受容体やイオンチャネルが存在することから、細胞感覚をつかさどる構造物として認識されるようになった。 例えば、嗅覚の受容体は樹状突起の繊毛に存在し、網膜の杆体や錐体の外節は、実は繊毛が変化したものである。 いくつかの受容体は他の細胞膜ドメインよりも繊毛膜に高度に充填されています。 また、繊毛の内部には、シグナル伝達の役割に関わる様々な分子が存在する。 繊毛の表面/体積比が高いため、繊毛内の分子反応は繊毛の外にある場合よりも強く、効率的である。 化学的なシグナル以外にも、一次繊毛は細胞外の液体の動きを感知し、メカノレセプターとして働くこともある。 べん毛
べん毛は繊毛に似ているが、長さは約150μmとずっと長く、わずかに太い。 細胞内では繊毛よりかなり数が少ない。 べん毛の主な働きは、細胞を動かすことである。 べん毛の運動は繊毛の運動とは異なり、細胞表面に対して垂直方向、つまりべん毛の長手軸の方向に運動する(平行ではない)。
構造
繊毛と鞭毛は、250 以上の異なるタンパク質を含む複雑な構造です。 両者とも、軸糸と呼ばれる中央の微小管組織と他の関連タンパク質を共有し、細胞膜によって制限されている(図3)。 また、繊毛・鞭毛の内部には、軸索以外に多くの可溶性分子が存在し、マトリックスを構成している。 軸索は9対の微小管で構成され、その周りにもう1対の中心微小管がある。 この構造は、(9×2)+2と書くことができる。 一次繊毛は、中央の一対の微小管を持たない。 中心対の微小管はそれぞれ13本のプロトフィラメントから構成されているが、周辺対の微小管は互いにいくつかのプロトフィラメントを共有している。 したがって、周辺部のペアはAおよびBの微小管によって形成される。

図3. 繊毛と鞭毛の主な分子構成要素。 一次繊毛では、中央のペアは存在しない。
軸糸の微小管編成は、タンパク質の足場がある結果である。 軸糸の微小管編成の維持に関与する軸糸の構成要素として、既に12種類のタンパク質が見つかっている。 近傍の周辺微小管対はネキシンによって互いに連結されている。 それぞれの対において、A微小管はタンパク質のスポークによって、中央の対や微小管を含む中央リングに接続されている。

図4. 脊髄上皮細胞の繊毛の超微細構造。 (9+2)x2とは、9組の周辺部と1組の中心部の微小管を意味する。
微小管は基底体から重合している(図3、図4)。 基底体は9本の3本鎖の微小管が円筒を形成している(遠心分離機に似ている)。 各3本鎖のうち、1本の微小管(A微小管)だけがプロトフィラメントを完全に持っているが、BとC微小管は互いにその一部を共有している。 基底体からAおよびB微小管が成長し、軸糸の周辺微小管を形成する。 基底体のすぐ上には、9対の周辺微小管を含む移行帯と呼ばれる繊毛の領域があり、中央の対はない。 移行帯の直後には基底板があり、ここから中央の一対の微小管が重合して軸糸を完成させる。 すべての微小管はプラス端が繊毛/鞭毛の先端に向かっている。 基底体の近位端(内側の微小管、つまりマイナス端)は、繊毛根状体と呼ばれる長いタンパク質繊維を通して細胞の細胞骨格に固定されている
繊毛・鞭毛には、軸糸の他にもコンパートメントがある。 膜には環境を感知するための多くの受容体やチャネルがあり、特に一次繊毛ではその数が多い。 内部の液相はマトリックスと呼ばれ、構造全体を組織的に保つのに役立ち、膜の受容体によって集められた情報を伝達する役割を担っています。 その他に、基部にある基底体、繊毛の先端部/鞭毛があり、微小管のプラス端を安定化させるタンパク質が含まれている。 運動
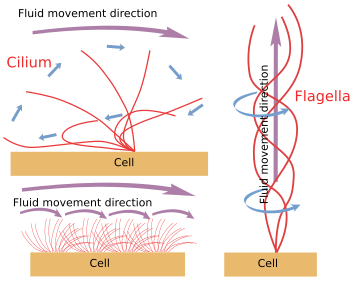
図6. 繊毛と鞭毛の運動に関するモデル。 これらは異なる流体移動方向を発生させる。
繊毛・鞭毛が機械的に細胞から切り離された場合、ATPストアが枯渇するまで動き続ける。 つまり、この運動機構は内在的なものである(図6)。 実は、この運動は、周辺部の微小管のペアが隣同士を滑らせることで生み出されている。 ネキシンとスポークタンパク質は、軸糸の乱れを防ぎつつ、このような動きを可能にする。 ダイニンは、滑走運動を担うモータータンパク質である。 エネルギーはATPによって供給される。 ダイニンは、球状部分を一方の周辺部の対のA微小管に、尾部を隣接する対のB微小管に固定されている。 その分子機構は細胞質ダイニンと似ているが、荷物を運ぶのではなく、微小管を移動させるという点である。 効率的な移動のためには、軸糸のダイニンの連携が必要である。 繊毛・鞭毛の内部にあるカルシウム波が、必要に応じてダイニンの活性化を調整し、運動頻度を変化させるのだろう。
5. 形成
分化の過程で、細胞は正常な生理機能に必要な繊毛と鞭毛をすべて作り出します。 すべてゼロから生成されなければならないことを意味する。 軸糸の微小管は基底体のA、B微小管から核生成されるので、繊毛・鞭毛1本につき1つの基底体が必要である。 複数の基底膜はどのように形成されるのか?
ヒトの多くの病態は、繊毛・鞭毛の欠陥の結果である。
ヒトの病理は、繊毛・鞭毛の欠陥の結果であり、繊毛症と呼ばれ、ランダムな側位、神経管の間違った閉鎖、多指症、嚢胞腎、肝臓・膵臓病理、網膜変性、肥満、認知障害などが含まれます。 繊毛:細胞のアンテナに同調する。 カレントバイオロジー。 16:R604-R614.
サティールP、クリステンセンST. 2007. 哺乳類繊毛の構造と機能の概要。 哺乳類繊毛の構造と機能の概要。 69:377-400.