Introduction
同時に現れる二つの視覚信号は、どちらかの信号だけが現れるよりも速く検知される(Hershenson,1962; Raab,1962)。 この冗長標的効果(RTE)の一部は、「競馬」モデルによって確率的に説明することができる。 すなわち、処理速度が試行ごとにランダムに変化する2つの刺激が並列かつ独立したチャネルで処理される場合、各提示で最も速い刺激がレースに勝ち、反応を誘発する検出閾値に達する。
しかし、累積頻度分布の分析により、2つの独立したチャネル間の競争馬モデルで説明できないRTEへの追加寄与が存在することが示されています。 Miller (1982) は、2つの信号が活性化プールで合計される「共活性化」モデルを提案した。 Miniussiら(1998)は、冗長な標的はP1およびN1事象関連脳電位の待ち時間を短くすることを示し、神経和が視覚経路の初期に生じることを示した。 また、半盲症患者の実験では、2つの刺激のうち一方を現象的に意識していなくてもRTEが発現することがあり、検出閾値到達の競馬では説明しきれないことが示されている(Marziら、1986; Tomaiuolo、1997;de Gelderら、2001;Lehら、2006a,b)。 実際、半球切除患者では、盲野の冗長な標的がRTEを生成するだけでなく、冗長な刺激がゲシュタルトパターンを完成させるとRTEが増強される(Georgy et al.、2016)。 半盲症におけるこれらの観察は,神経和が皮質下で起こり,一次視覚野に依存しないことを示唆している。
累積的な間接証拠は,RTEに寄与する神経和の基板として上丘(SC)を含意している。 しかし、直接的な証拠はまだ報告されていない。 本研究では、上丘で統合された視覚信号が伝達される神経経路を明らかにする。 SCは表層と深層から構成されている。 深層は眼球運動野から直接、あるいは基底核を経由して求心性神経を受け取る(Hikosaka and Wurtz, 1983; Pare and Wurtz, 1997; Sommer and Wurtz, 2000)。
視覚野を持たない神経疾患患者において神経和効果は実証されているので、小丘で和された視覚信号は網膜から網膜直腸路を経て小丘の表層に直接伝達されている可能性があります。
脊髄上膜を破壊するような病変は非常に稀であり,その場合,神経和は脊髄上膜の病変によって消失する可能性がある。
脳幹を損傷するような病変は非常に稀である。ここでは,視覚入力から脳幹を切り離す片側皮質下病変を持ち,RTEを示さなかった2名の患者について,単一症例研究を報告する。 1例は、視床のプルビナール核、吻側SCおよびその上腕の両方を損傷した病変を有していた。
材料と方法
2人の神経学者が、それぞれ年齢を合わせた対照群とともに、RTEを測定する実験に臨みました。 3年前の視床後部出血で内側滑車核、腹外側視床核、内嚢後縁が破壊され、損傷は前野と吻側SCを含む中脳背部に及んだ(図1)。確率的DTIトラクトグラフィー(Behrensら、2003、Behrensら、2007)により、SCの上腕部が破壊されたことが確認された(図2)。 図2)。REは左顔面および左腕の感覚を失い、左半身が麻痺していた。
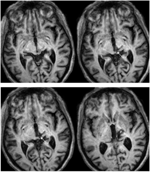
FIGURE 1. 患者REの高解像度(0.7 mm3)T1強調MRI。 右中脳背部と視床の病変を示す腹側(左上)から背側(右下)への軸位切片。
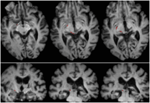
FIGURE 2.病巣の拡大(白い矢印で示される)。 T1強調軸方向スライス(上から腹側、背側)および冠状面スライス(下から前側、後側)に共登録された患者REの網膜直達路を示すトラクトグラフィ。 左半球に網膜直達路の走行を示す(赤)。 右半球では流線は描出されず、無傷の半球と比較すると、病変により右半球のSCの上腕が破壊されていることがわかる。 赤で示した流線は、FSL FDT(FMRIB Diffusion Toolbox; Behrens et al, 2003, Behrens et al, 2007; http://fsl.fmrib.ox.ac.uk/fsl/fslwiki) による確率的トラクトグラフィで生成した。 拡散強調エコー平面磁気共鳴画像は、32の等方的に分布する拡散エンコード方向(b=800)とベースライン(b=0)で、1.5 mm3の解像度で取得された。 繰り返し時間=2秒,エコー時間=35ms. 確率的トラクトグラフィでは、視交叉直後の各半球の視神経路に開始領域を手動でマーク(つまりシードマスクを描いた)し、SCに標的領域(ターゲットマスク)を描いた。
MLは検査時23歳であった。 7年前に交通事故で重度の外傷性脳損傷を受け,右被殻と中脳背部に出血を伴うびまん性軸索損傷があった。 病歴、神経学的検査、神経画像所見の詳細は(Poliva et al.、2015)により報告されている。 左は痙性、運動失調などの運動障害があるものの、自立性を取り戻していた。 主な障害は、左下小膠を含む脳幹聴覚路の損傷による重度の聴覚失認であった。 視力,視野に障害はなく,視覚無視,失認はなかった. 中脳機能障害による眼球運動徴候として,垂直注視時のマクロ矩形波ジャークと輻輳スパズム(上方より下方)がみられたが,眼瞼下垂や瞳孔異常はみられなかった. 高解像度MRIでは,前回出血部位の右被殻に嚢胞性空洞,右小脳下部に下小脳帯領域の小さな脳室周囲病変を認めた(図3). 左下小帯はほぼ完全に剥離し、最内側と尾側を残していた。 損傷は外側前胸部と中脳節に及び、上・下小柱のブラキアを破壊し、腹側には赤核に及んでいた。
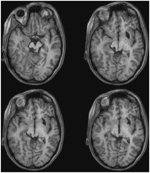
FIGURE 3. 患者MLの高解像度(0.7 mm3)T1強調MRI。 腹側(左上)から背側(右下)への軸方向スライスは、SCの腹側および外側における左下小丘と中脳節への損傷を示す。
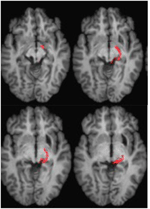
FIGURE 4.[1](図4)。 右半球の網膜-直腸路(赤)を示す患者MLのDTIトラクトグラフィは、腹側(左上)から背側(右下)までの軸平面でT1強調MR画像に共登録されている。 拡散強調エコー平面MR画像は2mm3の解像度で取得し、63の等方的に分布する拡散エンコード方向(b = 1000)とベースライン(b = 0)を設定した。 繰り返し時間=2秒、エコー時間=35ms。 トラクトグラフィーは図2のように実施した。
患者REの神経学的に健康な対照対象は、11人の高齢者(女性6人、男性5人、平均年齢:68.5、範囲59-73)である。 彼らは、バンガー大学のコミュニティ参加者パネルから募集された。 参加者は、既知の神経学的、心理学的、精神医学的、または認知障害を持たず、すべての参加者が正常/正視であった。
患者MLの神経学的に健康な対照被験者には、18~26歳のバンガー大学の大学生および大学院生11人(女性7人、男性4人)が含まれる。
装置と視聴条件
PCコンピュータ上で動作するPresentation®プログラミングソフトウェアが反応時間(RT)反応を記録し、Dellモニター(12.5″×25″、リフレッシュレート60Hz)上に提示する刺激を生成し、薄暗い条件で両眼視により参加者の前57cmに設置された。 頭部の安定性を確保するため、顎とヘッドレストを使用した。
刺激
実験刺激は、画面中央の小さな固定箱(0.2cm×0.2cm)の左右8°に配置された未記入の白いマーカー四角(1cm×1cm)で構成されていました。 標的刺激は、片方または両方のマーカーボックスが塗りつぶされ、白色の正方形が出現するものであった。
図5は、1回の試行の様子を示したものである。 マーカーボックスは実験中ずっと見えるままであった。 1750msの試行間隔後、固定箱が現れて試行が開始された。 250~750msのランダムな間隔(25ms刻み)の後、キーボードのスペースバーの単純なキープレス反応を必要とするターゲットが、一方または両方のマーカーボックスの位置に出現した。 ターゲットは、片方または両方のマーカーボックスが塗りつぶされた白色の正方形であった。 ターゲットは左側、右側、または両方に同じ確率でランダムに出現した。
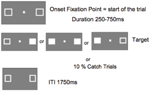
FIGURE 5.キャッチ試行(試行の10%)ではターゲットは現れなかった。
手順
各参加者は1回のセッションでテストされた。 画面上の指示を読んだ後、参加者は課題で提示されたすべての刺激(固定枠と3つの標的の提示それぞれ)の例を見せられ、スペースバーのキーを押してそれぞれの標的の認識を確認するよう求められた。
参加者がすべての指示を理解したことを口頭とキー操作で確認した後、練習試行(各ターゲット提示条件10回とターゲット不在の「キャッチ」試行3回)の提示が進められた。 練習試行終了後、画面上に以下のようなフィードバックが表示された。 (1)「正解」(2)「もっと速く反応して!」(3)「ターゲットが見えたときだけ反応して」。 練習セッションが終了し、参加者と実験者の両方が課題の指示を正しく理解したと確信した後、参加者は実験を開始するよう促された。 各実験ブロックの最初の30試行は、練習試行として除外された。 各ブロックは合計233試行で構成された。 右試行70回、左試行70回、両方70回、キャッチ試行23回。 ブロックの途中には短い休憩が与えられた。 患者MLと若年対照者は1つのブロックで試験を行った。 患者MLと若年対照者は、1つのブロックでテストを行った。患者REの年齢と状態による変動が大きいため、彼女は別々の日に2つのブロックでテストを行った。
解析
各コントロール群について、各条件(左ターゲット、右ターゲット、両側ターゲット)における各参加者のRT中央値は、キャッチ試行後の試行や、<100ms または >800ms の RT を除外して算出されました。 一対の標本t検定を行い、左右の片側標的に対する反応に非対称性がないことを確認し、片側標的に対する平均RTを算出した。 RTEは、片側試行のRTから両側試行のRTを引いて計算した。
各患者について、キャッチトライアルに続くトライアルと、RTが<100 msまたは>800 msのものは除外し、対のサンプルt-検定で対側と同側の片側ターゲットに対する中央値のRTを比較しました。 以下に報告するように、対側標的に対する反応のRTは、同側片側標的に対する反応よりも長かった。
各患者のRTEがそれぞれの対照群よりも確実に小さいかどうかを調べるために、各対照参加者のRTEを計算し、95%信頼区間の上限と下限を計算した。 各患者のRTEのZスコアは、それらの対照群と比較して計算され、Crawfordのt検定で統計的信頼性についてテストされた。
対のサンプルt検定のP値は、2尾、Crawford tの場合は1尾で報告された。
結果
患者のRE
対照参加者の場合。 図6は、両側標的のRTが、左側標的 (t = 6.8, p < 0.001, または右側標的 (t1,10) = 3.8, p = 0.003) に比べて短い (RTE) ことを示しています。 両側標的試行のRT中央値を片側標的のRT中央値の平均値から引き、各コントロール参加者の平均RTEを計算した。 対照群の平均RTEは24.3ms(SEM=2.75)であった。

FIGURE 6.平均RTEは、対照群の平均RTEは24.3ms(SEM=2.75)であった。 左視野(「左」)、右視野(「右」)、両視野に同時に現れる標的(「両方」)に対する患者REのRT中央値、および対照群参加者のRT中央値の平均値。
患者REのRT中央値は対照群よりも長く(図6)、3つの条件すべてにおいて対照群平均RTの95%CIの上限を超えていた(左=457ms、右=449ms、両方=426ms)。 また、対側(左)野の片側標的に対するRTは、同側(右)野の片側標的に対するRTよりも長かった(t = 6.0, p < 0.001).
重要なのは、RTEがなかったことです。両側標的のRT中央値(500ms)は、同側(右)視野の片側標的(494ms;t=1.141)より短くなりませんでした、p=ns(図6)です。 表1(上)は、患者REについて計算されたRTEが、彼女の対照群のRTEよりも有意に小さいことを示す。

TABLE 1. 患者RE(上)とML(下)の冗長標的効果、およびそれぞれの対照群の平均RTEと95%信頼区間(CI)、Zスコア、各患者とそれぞれの対照群を比較するクロフォードt検定。
患者 ML
対照群の参加者たち。 RTの中央値の平均は、左と右のターゲットへの応答について差がなかった(t = 0.86。 図7は、左(t = 4.7, p = 0.001)または右(t1,11)= 7.8, p < 0.001) 片側ターゲットよりも両側ターゲットで RT が短かった(RTE)ことを示す
 www.frontiersin.org
www.frontiersin.orgFIGURE 7.FIGE(図7)
両側標的試行の中央値RTは、各コントロール参加者のRTEを計算するために、左および右片側標的の中央値RTの平均から差し引かれた。
グループ平均RTEは33ms(SEM = 6.65)でした。
患者MLのRT中央値はコントロールよりも長く(図7)、3条件すべてにおいてコントロールグループ平均RTの95%CI上限を超えていました(左 = 322ms; 右 = 307ms; 両方 = 277ms )。 右側(対側)野の片側標的に対するRTは、左側(対側)野の片側標的に対するRTより長かった(t = 4.9, p = 0.001)。 RTEは見られなかった。 両側標的のRT(361ms)は同側視野の片側標的のRT(363ms)より短くはなかった。 表1(下)は、患者MLの計算したRTEが、彼女の対照群のRTEより有意に小さいことを示している。
考察
片半球のSC上腕が損傷した2人の患者において、対側視野と同側視野のいずれかにおける単一の標的、および両側標的を検出する単純RTを測定した。 そのうちの一人(若い女性)は、外傷性脳損傷による左半球の損傷であり、病変は下丘から中脳被蓋に及んでいたが、高解像度神経画像ではSC自体の損傷は明らかでなかった。 もう一人の患者(高齢女性)は、右半球の高血圧性出血によるもので、内側滑車と外側視床に広範な損傷を与え、吻側SCを含む中脳背部にまで及んだ。 両者とも対側視野の視覚的標的を検出する単純RTは同側視野に比べ遅かった.
これらの観察は,標的検出におけるSCの役割,より具体的にはSCへの視覚求心性の役割に光を当てるものであった。 ポズナーら(1980)は、検出を視覚信号に対する任意の反応(例えば、今回の実験のように指で単純にキーを押すこと)をする能力によって証明されるものと操作的に定義した。
Songら(2011)は、サルSCの不活性化が、視覚知覚や運動実行の単純な障害として容易に説明できない顕著な標的選択障害を引き起こすことを示し、それがより汎用的な優先度マップに寄与することを示唆した。 この実験では、サルに2つの刺激を対向する視野に提示し、どちらが先に現れたかをリーチング反応によって示させた。 1匹のサルは、最初に現れた刺激に向かって手を伸ばすように訓練され、1匹のサルは、2番目に現れた標的に向かって手を伸ばすように訓練された。 どちらの場合も、病変の反対側の視野にある標的を選択しないように強いバイアスがかかっていた。 この偏りは、2番目に現れた目標に手を伸ばしたサルにも見られたので、対側の目標の知覚の遅れに起因するものではないと思われる。 虹彩の不活性化は、どちらの刺激が先に出現したかの知覚判断に障害を生じさせなかった。
Zhaoping(2016)は最近、神経顕著性マップと優先度マップの区別を強調し、視蓋/SCから一次視覚野への知覚顕著性マップの進化的移行を主張した。
視索の上腕は、網膜から網膜直腸路を介して大脳皮質の表層に視覚信号を伝えるが、上腕を介して伝わる視覚求心線維の大部分は一次視覚野からの投射である。 上腕が障害されると、一次視覚野からの顕著な信号が前頭葉眼野を介して大脳皮質に伝達されるようになる。 しかし、上腕骨に障害がある患者のように、網膜や視覚野から小丘の表層に直接突起がない場合、小丘優先順位マップが活性化するまでに時間がかかり、結果として反対方向の視覚信号に対する反応が鈍くなることが予想されます。
冒頭で述べたように、確率的な競走馬モデルに基づく RTE への貢献は、両方のターゲットの検出と、2 つのターゲットを送信する独立したチャネルの効率が同等で、検出しきい値に達するレースの結果が試行ごとにランダムであるようなレースに依存しています。 つまり、同位信号と対位信号の間のレースの結果はランダムではなく、対位信号はレースに勝つよりも負ける可能性が高い。
にもかかわらず、半盲症患者の研究では、両方の標的の検出に依存しないRTEに寄与する神経和の成分も存在することが示されている (Marzi et al..)。 1986; Tomaiuolo, 1997; de Gelder et al., 2001)。
SavazziとMarzi(2004)は、神経学的に無傷の人とスプリットブレイン患者のいずれにおいても、短波長の色覚刺激では神経和が生じないことを示しました。 Lehら(2006b)は、半盲症患者において無彩色刺激ではRTEが起こるが、短波長の有彩色刺激を目標にするとRTEが起こらないことを報告している。 短波長(すなわち紫)の刺激は網膜のS錐体のみを活性化する。 S錐体からの入力を受けた網膜神経節細胞は、直接SCへも外側核の大細胞層にも投射しない(De Monasterio, 1978)。 一次視覚野からSCの上腕部を通ってSCに投射されるのは、大脳淋巴球求心性神経からの視覚信号のみを中継するため(Schiller et al,
したがって、スプリットブレイン患者、半盲症患者、および短波長刺激を用いた実験から、神経和がSCで起こるという収束した証拠が存在するのです。 しかし、短波長刺激は上腕骨を介してSCに投射する網膜神経節細胞を活性化しないが、小丘がそのような刺激に対して全く盲目であると結論づけることはできない。 サルの脊髄小胞における単一ユニットの記録は、短波長の刺激が脊髄小胞の表層における反応を活性化することを実証している。 さらに、トラクトグラフィーは、RTE(すなわち盲視)を有する半球切除患者において、盲視の証拠を示さない患者にはなく、SCが無傷の半球に接続していることを証明した(Leh et al, 2006a)。
したがって、今回の調査の結果は、空間的な総和が、その上腕を通して視覚信号の伝達に依存するSCで起こるという最初の直接的な証拠を提供する。
倫理声明
この研究はガイドラインに従い、英国バンゴーのNHS倫理委員会と英国バンガー大学の心理学倫理委員会のプロトコルに対する承認を受けて実施された。
著者貢献
MvK:研究の着想、実験の設計とプログラム、患者データの収集と分析、原稿執筆に貢献した。 KK:コントロールデータの収集、関連文献のレビュー、コントロールデータの分析、原稿執筆への貢献。
利益相反声明
著者らは、本研究が、潜在的な利益相反と解釈されうる商業的または金銭的関係がない状態で行われたことを宣言する。
Behrens, T, Woolrich, M. W., Jenkinson, M., Johansen-Berg, H., Nunes, R. G., Clare, S., et al.(2003)。 拡散強調MR画像における不確実性の特徴と伝播。 Magn. Reson. Med. 50, 1077-1088. doi: 10.1002/mrm.10609
PubMed Abstract | CrossRef Full Text | Google Scholar
Behrens, T. E., Berg, H. J., Jbabdi, S., Rushworth, M. F., and Woolrich, M. W. (2007). 複数の線維配向を用いた確率的拡散トラクトグラフィ:何を得ることができるのか? Neuroimage 34, 144–155.
Google Scholar
de Gelder, B., Pourtois, G., van Raamsdonk, M., Vroomen, J., and Weiskrantz, L. (2001). Unseen stimuli modulate conscious visual experience: evidence from interhemispheric summation. Neuroreport 12, 385–391. doi: 10.1097/00001756-200102120-00040
CrossRef Full Text | Google Scholar
De Monasterio, F. M. (1978). Properties of ganglion cells with atypical receptive-field organization in retina of macaques. J. Neurophysiol. 41, 1435–1449.
PubMed Abstract | Google Scholar
Fries, W. (1984). Cortical projections to the superior colliculus in the macaque monkey: a retrograde study using horseradish peroxidase. J. Comp. Neurol. 230, 55–76. doi: 10.1002/cne.902300106
PubMed Abstract | CrossRef Full Text | Google Scholar
Georgy, L., Celeghin, A., Marzi, C. A., Tamietto, M., and Ptito, A. (2016). 上丘は半球切除患者におけるゲシュタルト様刺激構成に敏感である。 Cortex 81, 151-161. doi: 10.1016/j.cortex.2016.04.018
PubMed Abstract | CrossRef Full Text | Google Scholar
Hershenson、M. (1962).。 このような場合、「感覚間ファシリテーションの尺度としての反応時間」、「感覚間ファシリテーションの尺度としての反応時間」が必要となる。 J. Exp. Psychol. 63, 289-293. doi: 10.1037/h0039516
CrossRef Full Text | Google Scholar
Hikosaka, O., and Wurtz, R. (1983). 彦坂翁、Wurtz, R. (1983). サル黒質網様体の視覚・眼球運動機能. サルの黒質網様体の視覚と眼球運動機能. J. Neurophysiol. 49, 1268-1284.
PubMed Abstract | Google Scholar
Leh, S. E., Johansen-Berg, H., and Ptito, A. (2006a). 無意識の視覚:拡散トラクトグラフィーを用いた盲視の神経細胞相関に関する新たな知見。 Brain 129(Pt 7), 1822–1832.
PubMed Abstract | Google Scholar
Leh, S. E., Mullen, K. T., and Ptito, A. (2006b). Absence of S-cone input in human blindsight following hemispherectomy. Eur. J. Neurosci. 24, 2954–2960.
PubMed Abstract | Google Scholar
Marzi, C., Tassinari, G., Aglioti, S., and Lutzemberger, L. (1986). Spatial summation across the vertical meridian in hemianopics: a test of blindsight. Neuropsychologia 24, 749–758. doi: 10.1016/0028-3932(86)90074-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Miller, J. (1982). Divided attention: evidence for coactivation with redundant signals. Cogn. Psychol. 14, 247–279. doi: 10.1016/0010-0285(82)90010-X
CrossRef Full Text | Google Scholar
Miniussi, C., Girelli, M., and Marzi, C. A. (1998). Neural site of the redundant target effect electrophysiological evidence. J. Cogn. Neurosci. 10, 216–230. doi: 10.1162/089892998562663
CrossRef Full Text | Google Scholar
Pare, M., and Wurtz, R. (1997). Monkey posterior parietal cortex neurons antidromically activated from superior colliculus. J. Neurophysiol. 78, 3493–3497.
PubMed Abstract | Google Scholar
Poliva, O., Bestelmeyer, P. E., Hall, M., Bultitude, J. H., Koller, K., and Rafal, R. D. (2015). ヒト聴覚野の機能マッピング:下丘の外傷による聴覚失認患者のfMRIによる検討。 Cogn. Behav. Neurol. 28, 160-180. doi: 10.1097/WNN.000000000072
PubMed Abstract | CrossRef Full Text | Google Scholar
Posner, M., Snyder, C., and Davidson, B. (1980)(1980). このような場合、「注意」と「信号の検出」が重要となる。 J. Exp. Psychol. Gen. 109, 160-174. doi: 10.1037/0096-3445.109.2.160
CrossRef Full Text | Google Scholar
Raab, D. H. (1962). 単純反応時間の統計的促進。 というものである。 N. Y. Acad. Sci. 24, 574-590. doi: 10.1111/j.2164-0947.1962.tb01433.x
CrossRef Full Text | Google Scholar
ロイター=ローレンツ、P、野沢、G、ガザニガ、M、S、ヒューズ、H・C・ (1995). このような場合、「redundant target effects in the bisected brain(冗長標的効果)」の時系列的な分析が必要となる。 J. Exp. Psychol. Hum. Percept. Perform. 21, 211-230. doi: 10.1037/0096-1523.21.2.211
PubMed Abstract | CrossRef Full Text | Google Scholar
Savazzi, S. and Marzi, C. A. (2004). 上丘は、健常者と脳梁の全切断または切断の患者の両方において、半球間神経和を維持する。 Neuropsychologia 42, 1608-1618. doi: 10.1016/j.neuropsychologia.2004.04.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Schiller, P.H., Malpeli, J.G. and Schein, S.J. (1979).のような。 J. Neurophysiol. 42, 1124-1133.
PubMed Abstract | Google Scholar
Sommer, M., and Wurtz, R. (2000). J. Neurophysiol. 83, 1979–2001.
PubMed Abstract | Google Scholar
Song, J. H., Rafal, R. D., and McPeek, R. M. (2011). PNAS Plus: deficits in reach target selection during inactivation of the midbrain superior colliculus. Proc. Natl. Acad. Sci. U.S.A. 108, E1433–E1440. doi: 10.1073/pnas.1109656108
PubMed Abstract | CrossRef Full Text
Tomaiuolo, F. (1997). Blindsight in hemispherectomized patients as revealed by spatial summation across the vertical meridian. Brain 120, 795–803. doi: 10.1093/brain/120.5.795
PubMed Abstract | CrossRef Full Text | Google Scholar
Zhaoping, L. (2016). From the optic tectum to the primary visual cortex: migration through evolution of the saliency map for exogenous attentional guidance. Curr. Opin. Neurobiol. 40, 94–102. doi: 10.1016/j.conb.2016.06.017
PubMed Abstract | CrossRef Full Text | Google Scholar