原理的に汎発的交配が破綻している家系や集団が存在する。 通常は地理的あるいは社会的に比較的隔離された小さな共同体である。 このようなコミュニティでは近親交配,つまり様々な順序(親-子,兄弟,いとこ,おじ-姪など)の血縁的な交配が広く行われていることがある。
近親交配が集団に与える影響の問題は、遺伝学の重要な問題である1。
ほとんどの場合、家系における近親交配は3世代で考慮されます。
家系における近親交配は、ほとんどの場合、3世代で考慮されます。
近親交配は近親交配係数の助けを借りて考慮されます。
近親交配係数は2つの対立遺伝子が起源によって同一である確率です。
ライト1、2が提供する家系図の近親交配係数計算の方法を考えてみましょう。
近親交配係数を求めるためにそれは必要である。
結婚する子孫について、共通の祖先を通してこれらの子孫をつなぐ家系図のすべての道を見つけること。
それぞれの道のステップ数(世代の交替)を数え上げること。
すると近親交配因子は次のように等しくなる。
もう一度、(1)の足し算の数は、共通の祖先を経て結婚に至る道の数に等しいことに注意しましょう。
図1の家系図の具体例を考えてみましょう。 この図では、四角が男性、丸が女性の個体である。 点線と同じ数字は、共通の祖先(男性または女性)を介して子孫の血縁結婚の接続家系図の可能な方法を指定します。 図1с、方法のいくつかのサイトでは、2つの数字1と2があります。
父と娘の結婚は近親相姦であり、図1а。 子孫(女性)は祖先(男性)と1回で結ばれる、つまりm=1なので近親交配因子が存在する。 桁は道の数を指定する。
結婚した子孫はそれぞれ2つの道を2歩ずつ相手と結ぶとすると、m=2であり、近親交配因子がある。 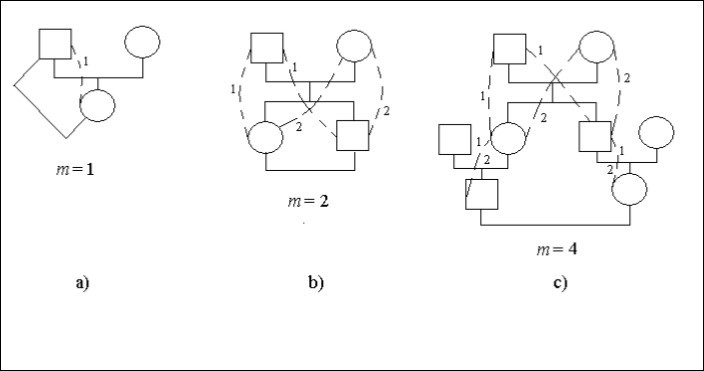
結婚した子孫はそれぞれ4歩ずつ、つまりm=4で2通り相手と繋がります。
Ft = 1/2.(2-4 + 2-4) = 1/16
式(1)は、以下の論理から容易に理解することができます。 女性から任意の祖先にηステップで道をリードしている場合は、この祖先との共通遺伝子のシェアは2-ηに等しくなります。 男から任意の祖先までがμステップで進むと、共通遺伝子のシェアは2-μに等しくなります。 男女が共通の祖先と結婚するとき、この祖先との共通遺伝子のシェアは2-ηになる。 2-μ=2-m ここでm=η+μは女性から男性が結婚するまでの共通祖先を通る総段階数である。 大きさ1/2 2-mは女性で偶然に選ばれた遺伝子が男性で偶然に選ばれた遺伝子と起源的に同一である確率を示す。
例えば、共通の祖先がη = μ = 5世代前に結婚していない場合、共通の遺伝子を持つ確率は等しくなります。 1/2 2-(5+5) = 1/2048 交配解析時のこの大きさは無視できる。 (表1)
表1.いくつかの近親婚の近親交配係数を追加で示した。
| 近親婚の種別 | |
| 近親婚 | いとこ | 1/32 |
| いとこのおじ – いとこ td niece | 1/32 |
| Third cousins | 1/64 |
The inbreeding factor Ft is the probability of two alleles in posterity are identical by origin i.e. occur from same gene of ancestor 1. Or, that too most, the inbreeding factor is a probability of the allele origin in posterity is consanguineous. Hence, the probability of the non-consanguineous origins of identical alleles in homologous chromosomes is equal 1 – Ft.
Let’s find total probability of the recessive homozygote aa occurrence (eventаа) at random mating for an autosomal genes 3:
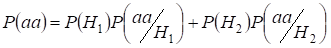 …….(2)
…….(2)
This formula follows from the following logic of mating: 相同な常染色体の同一対立遺伝子の起源が非血族的である場合(この仮説の確率は P(H1) = 1- Ft )、ホモ接合体が生じる(この事象の条件付確率は P(aa/H1) = q2)、または同一対立遺伝子の起源が血族的である場合(この仮説の確率は P(H2) = Ft )、ホモ接合体が生じる(この事象の条件付確率は P(aa/H2) = q )というものである。 後者の場合のホモ接合体aaの発生確率qは、対立遺伝子aが一方の常染色体にあれば、ホモ接合体の血縁関係の起源では必ず同じ対立遺伝子が相同の常染色体にあるはずであることから決定される。
したがって、劣性ホモ接合体の総発生確率は、
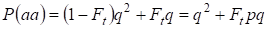 ……(3)
……(3)
ここで、p = 1 – qを考慮して、優勢対立遺伝子a頻度とする。
同様にランダム交配で優性ホモ接合体AAが出現する総確率を求める:
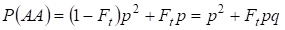 ……………………
……………………
ここで、p=1 – qを考慮すると、優性対立遺伝子aの頻度が存在することになる。 (4)
したがって、(3)と(4)によれば、近交系家系におけるホモ接合体(優性・劣性)の総頻度は2Ftpqで増加することになる。 したがってヘテロ接合体の頻度は同じ大きさでは減少し、すべての接合体頻度の和が1になるので、2pq – 2pqFt = 2pq(1-Ft)に等しくなる。
近親交配因子と(3)(4)を考慮すると、常染色体ゲノムの近親家系の次世代におけるハーディ-ワインバーグ則は、接合子の遺伝子型の分布を4と書き下す必要がある。
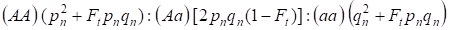 ……(5)
……(5)
ここでpnはn世代における優勢対立遺伝子A頻度、qnは劣勢対立遺伝子A頻度で、pn+qn=1である。
この比率(5)はライトの比率5でもある。 ライトの比率は近親交配後の家系において、汎発集団と比較して、Ft pnqnの家系では常染色体ホモ接合体の割合が増加し、(1-F1)では常染色体ヘテロ接合体の割合が減少するという効果が観察されていることを示すものである。
しかし、すでに1世代を経てハーディ-ワインバーグバランスが回復しており、家系にとって近親交配の別行動の重要性が小さいことを規定しています:
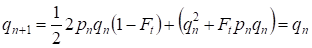 ……(6)
……(6)
明らかに、集団全体にとって近親交配という別行は家系にとってよりもさらに小さな重要性となるでしょう。
接合子の遺伝子型頻度のばらつきの効果は、性連鎖遺伝子についても同様である。
汎発家系におけるХ染色体に連なる遺伝子型の分布は、ハーディー・ワインベルグの法則により、1:
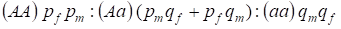 ……(7)
……(7)
優勢対立遺伝子Aの頻度は男性 pm、女性 pf で指定することにした。
交配時には、積 (pf + qf) (pm +qm) に従って、女性における遺伝子型の比率が存在する。
ここで、Х染色体に連なる娘の遺伝子について、無作為交配で劣性ホモ接合体aaが発生する総確率を式(2)のもとで求めてみよう。 事象の論理は常染色体の場合と同様であるが、(7)による非血縁対立遺伝子からの遺伝子型発生の条件付き確率はP(aa/H1)=qmnqfnに等しくなる。 近親対立遺伝子の場合、遺伝子型алаの発生する条件つき確率は P(aa/H2) = qf(n+1) に等しくなる。 娘たち、すなわち第n+1世代では、ホモ接合体aaの発生確率は、対立遺伝子aが一方のХ染色体にあれば、ホモ接合体の血族的起源では必ず同じ対立遺伝子が相同のХ染色体にあるはずであることによって決定されます。
したがって、劣性ホモ接合体の総発生確率は次のようになる:
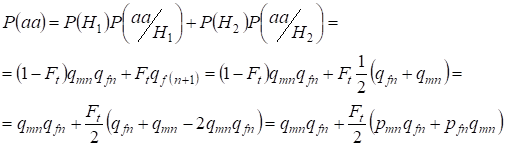 ……(8)
……(8)
ここでpmnはn世代における男性、pfn – 女性での優性対立遺伝子Aの頻度とする。 劣性対立遺伝子は、それに応じてqmnとqfn 。 また、qmn = 1 – qmn, pfn = 1 – qfnを用いる。
同様に、以下を用いて優性ホモ接合体の総発生確率を求める:
 ,
,
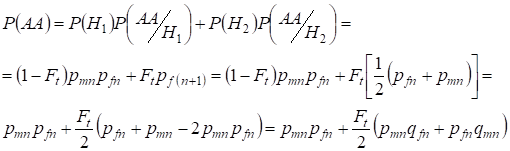 ……
……
……….
(9)
そしてヘテロ接合体 P(Aa) = 1- P(aa) – P(AA) 交配時の近交系家族樹のХ-染色体における遺伝子型の分布を求めよう:
 ……………….
……………….
…Х染色体におけるゲノム型分布は、交叉のときと同じである。…(10)
常染色体と同様に、性連鎖染色体における優性・劣性ホモ接合体頻度の近交系加算も同一であることが同時に証明される。
比率(10)の求め方は次のようになる。
まず、ランダムな血族的交配の機会におけるヘテロ接合体Aaの出現の総確率、すなわちP(Aa)を求めることにする。 この確率は次のような交配の論理から導かれる。 対立遺伝子の起源が非血族的である場合(この仮説の確率は P(H1) = 1 – Ft )、ヘテロ接合体 Aа も生じる(このイベントの条件付き確率は P(Aa/H1) = pmnqfn + pfnqmn.である)。 (7)式参照)、あるいは対立遺伝子の起源が血族的であり(この仮説の確率は P(H2) = Ft)、ヘテロ接合体 Aа が生じる(この事象の条件付確率は 0 P(Aa/H2) = 0 )ということです。
したがって、総確率の公式を用いて、ヘテロ接合体Aaの出現の総確率を求めると、
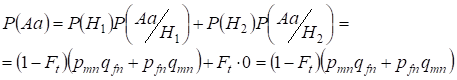 ………。…(11)
………。…(11)
優性AAと劣性aaのホモ接合体の頻度が同一に増加することにより、先に証明したようにヘテロ接合体Aaの頻度の減少が起こる。 例えば、劣性aaホモ接合体頻度の増加は等しい:
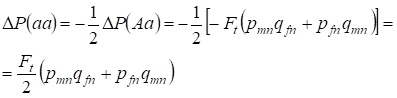 ……(12)
……(12)
ここで(7)と(11)によるパンミクティック家系に対するヘテロ接合体Aa頻度の減少が等しいとすると、以下のようになる。 ΔP(Aa) = – Ft (pmnqfn + pfnqmn) (11)と(12)を用いて近親交配の場合について(7)を更新すると、娘近親家系におけるХ-染色体の遺伝子型(10)の分布が求まる。
次の世代では、女性における劣性対立遺伝子aの頻度は等しく、
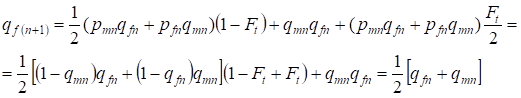 ……(13)
……(13)
これは近親交配のない場合の同じ頻度に完全に準拠し、つまりハーディー・ワインバーグ均衡の回復、より正確には均衡への接近があることになります。
比率(7)と(13)は、全体として近親交配を考慮した場合の家系における対立遺伝子頻度が変化しないことを示しています。 あるのは対立遺伝子の再分配だけである。 対立遺伝子の一部がヘテロ接合体から離れ、ホモ接合体に受け継がれるのである
。