The Atlantic cod is one of major predator in Barents Sea estimated to consume over 5 million tonnes of fish in 2017. 最近の論文(Holt et al. 2019)では、ロシアとノルウェーの科学者による33年間のタラの胃袋サンプリングを網羅したユニークなデータセットを使用して、この種の食事を調査しています。 この時系列は、これまでで最も包括的なタラの食餌データセットであり、タラが何を食べるのか、なぜそれがバレンツ海の生態系における捕食者-被食者と食物網の力学にとって重要なのか、生態学的に重要な質問に答える上で極めて重要である。
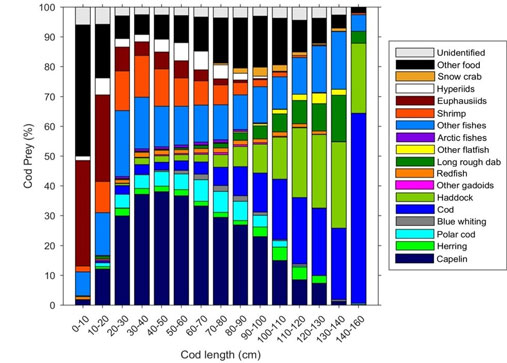
Figure 1.タラの食性。 サイズ(cm)の異なる大西洋マダラの食餌を、種/種群ごとの全餌生物重量に対する割合で表示。 データは1984年から2016年までの33年間の胃内容物の時系列データから来ている
種間の相互作用を理解することは、バレンツ海(投稿参照)のように、管理の生態系アプローチが望ましい海洋システムにおいて特に重要である。 経済的重要性(2017年の挑発的な報告漁獲量868 276トン:ICES 2018)と象徴的な地位のために、バレンツ海で最も研究されている種の1つは、北東北極タラ(Gadus morhua)である。 バレンツ海の漁業の主な対象であることに加え、タラはバレンツ海の食物網における著名な捕食者でもあります。 生態系の生物学的構成要素がどのように結びついているかを理解し、バレンツ海の資源開発を管理できるようにするためには、マダラの捕食動態に関する詳細な情報が必要である。 胃内容物データは、特定の時間・空間におけるある種の食性に関する包括的な情報を提供することができる。
1984年以来、PINRO (Knipovich Polar Research Institute of Marine Fisheries and Oceanography in Murmansk) とIMR (Institute for Marine Research in Bergen) は、バレンツ海でのデータを得るために共同している。 そのような協力関係の1つが、タラの胃袋の収集とその内容物の分析です。 長年にわたり、バレンツ海全体で380 948個の胃が採取され、毎年平均10 884個の胃が採取されるという素晴らしい結果が得られています。
Holtら(2019)は、これらのデータを使用して、バレンツ海におけるタラ食の変化、特に長期的な年間傾向、季節間の違い、タラと餌生物のサイズ関係、タラ食組成の発達中のシフトを調査しました。 この魚食傾向は、同種のタラ個体群よりも早い時期に確立されている。 共食いは、この種についてよく文書化されている (Bogstad et al. 1994; Dolgov et al. 1994; Uzars & Plikshs 2000; Neuenfeldt & Köster 2000; Yaragina et al. 2009; Pálsson & Björnsson 2011) は、すでに20cmのタラについて発見し、その強さはタラのサイズとともに増加した。 また、Holtらは、タラは自分の体長の33%が限界で、あらゆる大きさの獲物を食べることを発見した。
タラの食事は1984年から2016年にかけて大きく変化し、餌やタラの量や分布の変化と一致していた。 季節的な違いも観察され、冬の食事ではカラフトシシャモ(Mallotus villosus)が優勢であるのに対し、夏・秋にはタラ、極地タラ(Boreogadus saida)、その他の魚類がより優勢であった。 これは、この種の日和見的な捕食行動を裏付けるものである。
なぜこれらの知見が重要なのだろうか。
バレンツ海のタラは、バレンツ海で最も重要な捕食者と考えられている。 本研究により、タラの発達に伴う食餌組成のシフトと同様に、不釣り合いな捕食者-被食者のサイズ分布の証拠を提供することができた。 バレンツ海におけるマダラの資源量は近年増加しており、過去に例のないほど高い水準に達している。 そのため、タラの食性と資源量の関係は、重要な餌生物集団、特に餌の高い割合を占める遠洋魚類資源(ノルウェーの春産卵期のニシン Clupea harengus、カラフトマス…)とそれに依存する他の種への影響を強める可能性があります(投稿参照)。 このことは、バレンツ海の食物網の構造と生態系機能に影響を及ぼすと思われます。 この研究は、バレンツ海の生態系ダイナミクスを決定する栄養連鎖の理解に向けた重要なステップとなる。
Holt, R.E., Bogstad, B., Durant, J.M., Dolgov A.V., Ottersen, G. 2019年. Barents Sea cod (Gadus morhua) diet composition: long-term interannual, seasonal, and ontogenetic patterns. ICES Journal of Marine Science DOI: 10.1093/icesjms/fsz082
Bogstad, B., Lilly, G.R., Mehl, S., Pálsson, Ó.K. Stefánsson, G. 1994年. ICESジャーナル・オブ・マリンサイエンス。 198: 576-599.
Dolgov, A.V., Korzhev, V.L., Tretyak, V.L. 1994. バレンツ海におけるアークト・ノルウェー産マダラの加入形成における共食いとその重要性。 121-128.
ICES. 2018. 北極漁業作業部会(AFWG)報告書、2018年4月18-24日、イタリア、イスプラ。 ices cm 2018/acom:06. 859頁
Neuenfeldt,S.、Köster,F.W.2000. Trophodynamic control on recruitment success in Baltic cod: the influence of cannibalism. ICESジャーナル・オブ・マリンサイエンス. 57: 300-309.
Pálsson, Ó.K., Björnsson, H. 2011. このような状況下において、「鯨類と鯨類との関わり」は、「鯨類と鯨類との関わり」、「鯨類と鯨類との関わり」、「鯨類と鯨類との関わり」の3つに分類される。 ICES Journal of Marine Science. 68: 1488-1499.
Uzars, D., Plikshs, M. 2000. ICES Journal of Marine Science. 57: 324-329
Yaragina、N.A.、Bogstad、B.、Kovalev、Y.A. 2009. 北東北のタラ(Gadus morhua)における1947年から2006年までのカニバリズムの変動性。 Marine Biology Research. 5: 75-85