Introducere
Două semnale vizuale care apar simultan sunt detectate mai rapid decât oricare dintre semnalele care apar singure (Hershenson, 1962; Raab, 1962). O parte din acest efect de țintă redundantă (RTE) poate fi explicat în mod stocastic printr-un model „horse-race”. Adică, dacă doi stimuli sunt procesați în canale paralele și independente pentru care viteza de procesare variază aleatoriu de la o încercare la alta, la fiecare prezentare, cel mai rapid stimul câștigă cursa și atinge pragul de detecție pentru a declanșa răspunsul; prin urmare, în medie, este mai probabil ca doi stimuli să producă un răspuns mai rapid decât timpul mediu de răspuns la un singur stimul procesat în oricare dintre cele două canale.
Cu toate acestea, analizele distribuțiilor de frecvențe cumulative au arătat că există o contribuție suplimentară la RTE care nu poate fi luată în considerare de un model al cursei de cai între două canale independente. Miller (1982) a propus un model de „co-activare” în care cele două semnale sunt însumate într-un grup de activare. Miniussi et al. (1998) au arătat că țintele redundante au produs latențe mai scurte pentru potențialele cerebrale legate de evenimente P1 și N1, ceea ce indică faptul că însumarea neuronală are loc devreme în calea vizuală. Experimentele la pacienții hemianopici au arătat, de asemenea, că RTE nu poate fi pe deplin explicată printr-o cursă de cai pentru a atinge pragul de detecție, deoarece o RTE se poate manifesta chiar și în absența conștientizării fenomenale a unuia dintre cei doi stimuli (Marzi et al., 1986; Tomaiuolo, 1997; de Gelder et al., 2001; Leh et al., 2006a,b). Într-adevăr, la pacienții cu hemisferectomie, o țintă redundantă în câmpul orb nu numai că generează o RTE, dar RTE este augmentată atunci când stimulul redundant completează un model gestalt (Georgy et al., 2016). Aceste observații la hemianopici sugerează că însumarea neuronală are loc la nivel subcortical și nu depinde de cortexul vizual primar.
Dovezile indirecte cumulative au implicat coliculul superior (SC) ca fiind substratul pentru însumarea neuronală care contribuie la RTE. Cu toate acestea, nu au fost încă raportate dovezi directe. Cercetarea actuală investighează calea neuronală care transmite semnalele vizuale care sunt integrate în SC. SC este format din straturi superficiale și profunde. Straturile profunde ale coliculului primesc aferențe, atât direct, cât și prin ganglionii bazali, de la cortexul oculomotor (Hikosaka și Wurtz, 1983; Pare și Wurtz, 1997; Sommer și Wurtz, 2000). Cu toate acestea, aferențele vizuale către straturile superficiale ale coliculului sunt transmise atât de la retină (prin intermediul tractului retino-tectal), cât și de la cortexul vizual timpuriu (Fries, 1984), prin intermediul brațului SC.
Din moment ce efectele de însumare neuronală au fost demonstrate la pacienții neurologici lipsiți de cortex vizual, semnalele vizuale însumate în colicul ar putea fi transmise direct de la retină la straturile superficiale ale coliculului prin intermediul tractului retino-tectal. Dacă acesta este cazul, însumarea neuronală ar putea fi abolită prin leziuni ale brahiului SC.
Lesiunile care perturbă brahiul SC sunt foarte rare. Aici raportăm studii de caz unic a doi pacienți cu leziuni subcorticale unilaterale care au dezaferentat SC de intrarea vizuală și care nu au manifestat o RTE. Un pacient a avut o leziune care a afectat atât nucleul pulvinar al talamusului, SC rostral, cât și brahiul acestuia. La celălalt pacient, cu leziuni la nivelul mezencefalului dorsal în urma unei leziuni cerebrale traumatice, SC părea să fie intact, dar brahiul său era perturbat.
Materiale și metode
Participanți
Doi pacienți neurologici, fiecare cu propriul grup de control, comparat ca vârstă, au fost testați în cadrul unui experiment pentru a măsura RTE.
RE avea 71 de ani la momentul testării. O hemoragie talamică posterioară cu 3 ani mai devreme a distrus pulvinarul medial, nucleii talamici ventro-laterali și membrul posterior al capsulei interne, cu leziuni care se extind în mezencefalul dorsal, inclusiv pretectul și SC rostral (Figura 1) Tractografia probabilistică DTI (Behrens et al., 2003, Behrens et al., 2007) a confirmat faptul că leziunea a distrus brahiul din SC (Figura 2). RE era paralizată pe partea stângă a corpului, cu pierderea sensibilității la nivelul feței și brațului stâng. Avea mișcări anormale ale ochilor, inclusiv retracția pleoapelor, mișcări paralizate ale ochilor pe verticală și convergență afectată, care au fost o consecință a extinderii leziunii în mezencefalul dorsal.
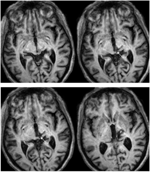
FIGURA 1. RMN de înaltă rezoluție (0,7 mm3) cu ponderare T1 a pacientei RE. Felii axiale de la ventral (stânga sus) la dorsal (dreapta jos) care arată leziunea din mezencefalul dorsal drept și talamusul. Extinderea leziunii în coliculul superior rostral (SC) arătată prin săgeți albe.
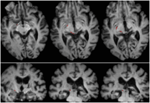
FIGURA 2. Tractografie care demonstrează tractul retinotectal la pacientul RE co-înregistrat la felii axiale ponderate T1 (Sus, de la ventral la dorsal) și la felii coronale (Jos, de la anterior la posterior). Cursul tractului retinotectal este prezentat în emisfera stângă (roșu). În emisfera dreaptă nu a fost trasată nicio linie de traiect și, prin comparație cu emisfera intactă, se poate observa că leziunea a distrus brahiul SC în emisfera dreaptă. Linia streamline prezentată în roșu a fost generată cu tractografia probabilistică utilizând FSL FDT (FMRIB Diffusion Toolbox; Behrens et al., 2003, Behrens et al., 2007; http://fsl.fmrib.ox.ac.uk/fsl/fslwiki). Imaginile de rezonanță magnetică eco-planare ponderate prin difuzie au fost achiziționate la o rezoluție de 1,5 mm3 cu 32 de direcții de codificare a difuziei distribuite izotropic (b = 800) și o linie de bază (b = 0). Timpul de repetiție = 2 s, iar timpul de ecou = 35 ms. Pentru tractografia probabilistică, am marcat manual regiunea de pornire (adică am desenat măștile de semințe) pe tractul optic al fiecărei emisfere imediat după chiasmă și o regiune țintă (masca țintă) pe SC.
ML avea 23 de ani la momentul testării. Ea suferise un traumatism cranio-cerebral sever în urma unui accident rutier cu 7 ani mai devreme, care a avut ca rezultat o leziune axonală difuză cu hemoragie în putamenul drept și în mezencefalul dorsal. Detalii despre istoricul ei, examinarea neurologică și rezultatele neuroimagistice au fost raportate de (Poliva et al., 2015). Deși a rămas cu unele deficiențe motorii, inclusiv spasticitate și ataxie, ea și-a recăpătat independența. Principala ei dizabilitate a fost o agnosie auditivă severă datorată deteriorării căilor auditive ale trunchiului cerebral, inclusiv a coliculului inferior stâng. Acuitatea vizuală și câmpurile vizuale erau neafectate și nu existau neglijență vizuală sau extincție vizuală. Semnele oculomotorii ale disfuncției mezencefalice au inclus sacadări ale undelor macro-pătrate și spasm de convergență la privirea verticală (mai mult în jos decât în sus.) Nu a existat ptoză sau anomalie pupilară. Rezonanța magnetică de înaltă rezoluție a arătat o cavitate chistică în putamenul drept la locul hemoragiei anterioare și o mică leziune periventriculară în partea inferioară a ponsului drept, în regiunea pedunculului cerebelos inferior (figura 3). Ea a avut o avulsie aproape completă a coliculului inferior stâng, cruțând doar părțile sale cele mai mediale și caudale. Leziunile s-au extins în pretectul lateral și în tegmentul mezencefalic, distrugând brahiile coliculilor superior și inferior, și ventral în nucleul roșu. Tractografia probabilistică DTI a confirmat faptul că leziunea a distrus brahiile SC (figura 4).
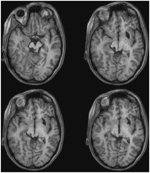
FIGURA 3. RMN de înaltă rezoluție (0,7 mm3) cu ponderare T1 la pacientul ML. Felii axiale de la ventral (stânga sus) la dorsal (dreapta jos) care arată leziuni ale coliculului inferior stâng și ale tegmentului mezencefalic ventral și lateral față de SC.
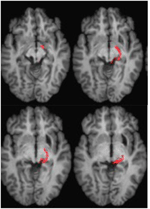
FIGURA 4. Tractografia DTI la pacientul ML care arată tractul retino-tectal din emisfera dreaptă (roșu) co-înregistrat la imaginile RM ponderate T1 în plan axial de la ventral (stânga sus) la dorsal (dreapta jos). Imaginile de rezonanță magnetică eco-planare ponderate prin difuzie au fost achiziționate la o rezoluție de 2 mm3 cu 63 de direcții de codificare a difuziei distribuite izotropic (b = 1000) și o linie de bază (b = 0). Timpul de repetiție = 2 s, iar timpul de ecou = 35 ms. Tractografia a fost implementată așa cum este descrisă în figura 2.
Subiectul de control neurologic sănătos pentru pacientul RE a inclus 11 adulți în vârstă (șase femei și cinci bărbați, vârsta medie: 68,5, interval 59-73). Aceștia au fost recrutați din panoul de participanți din comunitatea Universității Bangor. Participanții nu aveau nicio afecțiune neurologică, psihologică, psihiatrică sau cognitivă cunoscută și toți participanții aveau o vedere normală/corectată spre normal. Participanții au primit o plată de 6 lire sterline pentru participarea lor.
Subiecții de control sănătoși din punct de vedere neurologic pentru pacientul ML au inclus 11 studenți și postuniversitari de la Universitatea Bangor cu vârste cuprinse între 18 și 26 de ani (șapte femei și patru bărbați).
Aparatură și condiții de vizualizare
Software-ul de programare Presentation® care rulează pe un computer PC a înregistrat răspunsurile timpului de reacție (RT) și a generat stimuli care au fost prezentați pe un monitor Dell (12,5″ × 25″, rata de reîmprospătare 60 Hz.) plasat la nivelul ochilor la 57 cm în fața participanților cu vizualizare binoculară în condiții de lumină slabă. O bărbie și o tetieră au fost utilizate pentru a asigura stabilitatea capului. Răspunsurile participanților au fost înregistrate prin apăsarea barei de spațiu pe o tastatură.
Stimuli
Stimulii experimentali au constat în pătrate de marker alb necompletate (1 cm × 1 cm) poziționate la 8° la stânga și la dreapta unei mici cutii de fixare (0,2 cm × 0,2 cm) în centrul ecranului. Stimulii țintă au constat în umplerea uneia sau a ambelor casete de marcare periferice pentru a produce un pătrat alb solid. Toți stimulii au fost albi prezentați pe un fundal gri.
Figura 5 prezintă secvența unui singur proces. Casetele de marcare au rămas vizibile pe tot parcursul experimentului. După un interval între încercări de 1750 ms, caseta de fixare a apărut pentru a începe încercarea. După un interval aleatoriu variind de la 250 la 750 ms (în trepte de 25 ms), o țintă care necesita un răspuns prin simpla apăsare a unei taste pe bara de spațiu de la tastatură, a apărut la locația uneia sau a ambelor cutii de marcare. Ținta a fost un pătrat alb umplut, generat prin umplerea uneia sau a ambelor cutii de marcaj. În mod aleatoriu și cu o probabilitate egală, țintele au apărut în stânga, în dreapta sau în ambele simultan. Nicio țintă nu a apărut în încercările de captură (10% din încercări).
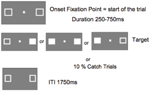
FIGURA 5. Secvența de afișare a unui singur proces (ITI, intervalul inter-trial între răspunsul participantului și apariția punctului de fixare care începe următorul proces).
Procedură
Care participant a fost testat într-o singură sesiune. După ce au citit instrucțiunile de pe ecran, participanților li s-au arătat exemple ale tuturor stimulilor care au fost prezentați în cadrul sarcinii (caseta de fixare și fiecare dintre cele trei prezentări ale țintei) și li s-a cerut să confirme recunoașterea fiecărei ținte prin apăsarea tastei space-bar. Instrucțiunile au fost de a menține fixarea pe caseta centrală pe durata fiecărui proces și de a răspunde cât mai repede posibil prin apăsarea barei de spațiu de la tastatură cu mâna dreaptă de îndată ce apărea o țintă.
După ce participantul a confirmat verbal și prin apăsarea tastelor că toate instrucțiunile au fost înțelese, s-a trecut la prezentarea încercărilor de practică (10 din fiecare condiție de prezentare a țintei și 3 încercări de „captură” fără țintă). Feedback-ul de pe ecran a fost prezentat în timpul finalizării încercărilor de practică, incluzând următoarele afirmații: (1) „Corect”, (2) „Încercați să răspundeți mai repede!” și (3) „Răspundeți numai dacă vedeți o țintă”. Odată ce sesiunea de practică a fost finalizată și atât participantul, cât și experimentatorul au fost siguri că instrucțiunile sarcinii au fost înțelese corect, participantul a fost invitat să înceapă experimentul. Primele 30 de încercări din fiecare bloc experimental au fost excluse ca încercări de practică. Fiecare bloc a constat dintr-un total de 233 de încercări: 70 de încercări la dreapta, 70 la stânga, 70 la ambele și 23 de încercări de „captură”. O scurtă pauză a fost acordată la jumătatea blocului. Pacientul ML și martorii mai tineri au fost testați într-un singur bloc. Din cauza variabilității legate de vârsta și afecțiunea pacientei RE, aceasta a fost testată în două blocuri în zile separate; iar participanții de control de vârstă corespunzătoare au fost, de asemenea, testați în două sesiuni într-o singură zi.
Analize
Pentru fiecare grup de control, RT median pentru fiecare participant în fiecare condiție (țintă stângă, țintă dreaptă, ținte bilaterale) a fost calculat după excluderea încercărilor care au urmat încercărilor de prindere și a celor cu RT-uri de <100 ms sau >800 ms. S-a efectuat un test t cu eșantion împerecheat pentru a confirma că nu a existat asimetrie pentru răspunsurile la ținte unilaterale drepte și stângi și s-a calculat un RT mediu pentru ținte unilaterale. RTE a fost calculat prin scăderea RT pentru încercări bilaterale din RT pentru încercări unilaterale. Un test t cu eșantion împerecheat a comparat unilateral cu bilateral pentru a stabili dacă RTE a fost fiabil din punct de vedere statistic.
Pentru fiecare pacient, după excluderea încercărilor care au urmat încercărilor de prindere și a celor cu RT de <100 ms sau >800 ms, un test t cu eșantion împerecheat a comparat mediile RT pentru țintele unilaterale contralaterale și ipsilaterale. După cum se raportează mai jos, RT-urile pentru răspunsurile la țintele contralesionale au fost mai lungi decât pentru țintele unilaterale ipsilare. Pentru a determina dacă prezența unei ținte contralesionale a generat un RTE fiabil din punct de vedere statistic, un test t pe eșantioane împerecheate a comparat mediile RT pentru încercările cu ținte bilaterale cu încercările cu ținte unilaterale ipsilesionale.
Pentru a testa dacă RTE a fost mai mic în mod fiabil pentru fiecare pacient decât pentru grupul de control respectiv, RTE a fost calculat pentru fiecare participant de control și au fost calculate limitele superioare și inferioare ale intervalelor de încredere (IC) de 95%. S-a calculat scorul Z pentru RTE al fiecărui pacient în raport cu grupul lor de control și s-a testat fiabilitatea statistică cu testul t al lui Crawford.
Valorile P pentru testele t cu eșantioane împerecheate au fost raportate pentru două cozi, iar pentru testul t al lui Crawford, cu o coadă.
Rezultate
RTE pacienți
Participanți la control: Media mediilor RT nu a fost diferită pentru răspunsurile din stânga și din dreapta (t = 0,6.) Figura 6 arată că RT au fost mai scurte (RTE) pentru țintele bilaterale decât pentru țintele unilaterale din stânga (t = 6,8, p < 0,001, sau din dreapta (t1,10) = 3,8, p = 0,003). Mediana RT pentru încercările cu ținte bilaterale a fost scăzută din media medianei RT pentru țintele unilaterale pentru a calcula un RTE mediu pentru fiecare participant de control. RTE mediu al grupului de control a fost de 24,3 ms (SEM = 2,75).

FIGURA 6. Mediile RT pentru pacientul RE și media mediilor RT pentru participanții din grupul de control pentru țintele care apar în câmpul vizual stâng („Left”), în câmpul vizual drept („Right”) și simultan în ambele câmpuri vizuale („Both”). Barele de eroare pentru participanții de control denotă erori standard.
Patientul RE a avut RT-uri mediane mai lungi decât controalele (Figura 6), și au fost peste limita superioară a IC 95% a RT-urilor medii ale grupului de control pentru toate cele trei condiții (stânga = 457 ms; dreapta = 449 ms; ambele = 426 ms). RT-urile pentru țintele unilaterale în câmpul contralesional (stânga) au fost mai lungi decât pentru țintele unilaterale în câmpul ipsilesional (dreapta) (t = 6,0, p < 0,001).
Important, nu a existat un RTE: RT median pentru țintele bilaterale (500 ms) nu a fost mai scurt decât pentru țintele unilaterale în câmpul vizual ipsilesional (drept) (494 ms; t = 1,141), p = ns (Figura 6). Tabelul 1 (sus) arată că RTE calculat pentru pacienta RE a fost semnificativ mai mic decât RTE pentru grupul ei de control.

TABEL 1. Efectul țintă redundant pentru pacienții RE (sus) și ML (jos) și RTE mediu pentru grupurile lor de control respective, cu intervale de încredere (IC) de 95%, scoruri Z și testul t Crawford care compară fiecare pacientă cu grupul ei de control respectiv.
Pacient ML
Participanți de control: Media mediilor RT nu a fost diferită pentru răspunsurile la țintele din stânga și din dreapta (t = 0,86. Figura 7 arată că RT-urile au fost mai scurte (RTE) pentru țintele bilaterale decât pentru țintele unilaterale din stânga (t = 4,7, p = 0,001, sau din dreapta (t1,11) = 7,8, p < 0,001).

FIGURA 7. Mediile RT pentru pacientul ML și media mediilor RT pentru participanții din grupul de control pentru țintele care apar în câmpul vizual stâng, în câmpul vizual drept și simultan în ambele câmpuri vizuale. Barele de eroare pentru participanții din grupul de control denotă erori standard.
RT mediana pentru încercările cu ținte bilaterale a fost scăzută din media medianei RT pentru țintele unilaterale din stânga și din dreapta pentru a calcula un RTE pentru fiecare participant de control.
RTE-ul mediu al grupului a fost de 33 ms (SEM = 6.65).
RTE-urile mediane ale pacientului ML au fost mai lungi decât cele ale controalelor (Figura 7) și au fost peste limita superioară a IC 95% a RT-urilor medii ale grupului de control pentru toate cele trei condiții (stânga = 322 ms; dreapta = 307 ms; ambele = 277 ms). RT-urile pentru țintele unilaterale din câmpul drept (contralesional) au fost mai lungi decât pentru țintele unilaterale din câmpul stâng (ipsilesional) (t = 4,9, p = 0,001). Nu a existat niciun RTE. RT pentru țintele bilaterale (361 ms) nu au fost mai scurte decât pentru țintele unilaterale din câmpul vizual ipsilesional (stâng) (363 ms). Tabelul 1 (partea de jos) arată că RTE calculat pentru pacienta ML a fost semnificativ mai mic decât RTE pentru grupul ei de control.
Discuție
S-au măsurat RT-uri simple pentru a detecta ținte unice în câmpul contralesional sau ipsilesional, și ținte bilaterale, la doi pacienți la care brahiul SC a fost afectat într-o emisferă. La unul dintre pacienți (o femeie tânără), leziunea s-a produs în emisfera stângă și s-a datorat unei leziuni cerebrale traumatice; și, în timp ce leziunea a implicat coliculul inferior și s-a extins în tegmentul mezencefalic, afectarea SC în sine nu a fost evidentă cu ajutorul neuroimagisticii de înaltă rezoluție. La celălalt pacient (o femeie mai în vârstă), leziunea a fost în emisfera dreaptă și a fost cauzată de o hemoragie hipertensivă care a provocat, de asemenea, leziuni extinse la nivelul pulvinarului medial și al talamusului lateral și s-a extins în mezencefalul dorsal, inclusiv în SC rostral. La ambii pacienți, RT simplu pentru detectarea țintelor vizuale în câmpul contralesional a fost încetinit în comparație cu câmpul ipsilesional. Niciunul dintre pacienți nu a prezentat un efect de însumare spațială (RTE) pentru detectarea țintelor atunci când stimulii au fost prezentați în ambele câmpuri vizuale.
Aceste observații aruncă lumină asupra rolului SC în detectarea țintelor și, mai precis, asupra rolului aferențelor vizuale către SC. Posner și colab. (1980) au definit operațional detecția ca fiind evidențiată prin capacitatea de a da un răspuns arbitrar la un semnal vizual (de exemplu, o simplă apăsare a unei taste cu un deget, ca în experimentul actual). Aceștia au presupus că detectarea necesită o alocare a atenției pentru a selecta ținta pentru procesare într-un sistem cu capacitate limitată care îi acordă prioritate pentru a fi acționat.
Song et al. (2011) au arătat că inactivarea SC la maimuțe cauzează deficite izbitoare de selecție a țintei care nu pot fi explicate cu ușurință ca o simplă afectare a percepției vizuale sau a execuției motorii și au sugerat că aceasta contribuie la o hartă de prioritate cu scop mai general. În acel experiment, maimuțelor li s-au prezentat doi stimuli în câmpuri vizuale opuse și trebuiau să indice, cu un răspuns de atingere, care a apărut primul. O maimuță a fost antrenată să se îndrepte spre stimulul care a apărut primul; și o maimuță a fost antrenată să se îndrepte spre ținta care a apărut a doua. În ambele cazuri, a existat o prejudecată puternică împotriva selectării țintei care se afla în câmpul vizual contralateral la leziune. Deoarece aceeași prejudecată a fost observată la maimuța care a întins mâna spre a doua țintă care a apărut, prejudecata nu a putut fi atribuită unei întârzieri în perceperea țintei contralesionale. Inactivarea coliculară nu a cauzat o afectare a judecății perceptuale a stimulului care a apărut primul. Mai degrabă, stimulii din câmpul vizual contralateral coliculului inactivat au fost dezavantajați în a fi prioritizați pentru acțiune.
Zhaoping (2016) a evidențiat recent o distincție între hărțile neuronale de saliență și hărțile de prioritate și a argumentat pentru o migrare evolutivă a unei hărți de saliență perceptivă de la tectul optic/SC la cortexul vizual primar. Ea a propus că semnalele de saliență pot fi transmise unei hărți de prioritate în SC fără a proiecta proprietatea de reglare a trăsăturilor.
Brachiul din SC transmite semnale vizuale aferente către straturile superficiale ale coliculului de la retină prin intermediul tractului retino-tectal; dar majoritatea fibrelor aferente vizuale transmise prin brahium sunt proiecții din cortexul vizual primar. În cazul în care brahiul este perturbat, semnalele de saliență de la cortexul vizual primar pot fi retransmise către colicul prin intermediul câmpurilor oculare frontale. Dar în absența proiecțiilor directe către straturile superficiale ale coliculului dinspre retină sau dinspre cortexul vizual, așa cum presupunem că este cazul pacienților cu leziuni ale brahiului, este nevoie de mai mult timp pentru activarea hărții de prioritate coliculară, ceea ce duce la răspunsuri mai lente la semnalele vizuale contralesionale.
Așa cum s-a menționat în introducere, contribuția la RTE care se bazează pe un model probabilistic al calului de curse depinde de detectarea ambelor ținte; și de o cursă în care eficiența canalelor independente în care sunt transmise cele două ținte este echivalentă, astfel încât rezultatul cursei pentru atingerea pragului de detecție este aleatoriu de la un proces la altul. Deoarece pragul de detecție pentru semnalele contralesionale este mai mare la pacienții raportați aici, țintele redundante nu ar putea beneficia pe baza unei curse de cai stocastice; adică, rezultatul cursei dintre semnalele ipsilesionale și contralesionale nu este aleatoriu – semnalele contralesionale au mai multe șanse să piardă cursa decât să o câștige.
Cu toate acestea, cercetările la pacienții hemianopici au arătat că există, de asemenea, o componentă de însumare neuronală care contribuie la RTE, care nu depinde de detectarea ambelor ținte (Marzi et al., 1986; Tomaiuolo, 1997; de Gelder et al., 2001). Deoarece s-a demonstrat că pacienții comisurotomizați au o RTE (Marzi et al., 1986; Reuter-Lorenz et al., 1995), s-a argumentat că integrarea interhemisferică a semnalelor de-a lungul meridianului vertical trebuie să aibă loc la nivel subcortical.
Savazzi și Marzi (2004) au arătat că însumarea neuronală nu a avut loc cu stimuli țintă cromatici de lungime de undă scurtă nici la persoanele intacte din punct de vedere neurologic, nici la pacienții cu creier divizat. Leh et al. (2006b) au raportat că, în timp ce o RTE a avut loc cu stimuli acromatici la unii pacienți hemianopici hemisferectomizați, nu a existat o RTE la acești pacienți atunci când stimulii cromatici de lungime de undă scurtă au fost utilizați ca ținte. Stimulii cu lungime de undă scurtă (de exemplu, violet) activează numai conurile S din retină. Celulele ganglionare ale retinei care primesc intrări de la conurile S nu se proiectează nici direct către SC, nici către straturile magnocelulare ale nucleului geniculat lateral (De Monasterio, 1978). Deoarece proiecțiile de la cortexul vizual primar către SC prin brațul SC retransmit doar semnale vizuale de la aferențele geniculostriate magnocelulare (Schiller și colab…, 1979), eșecul stimulilor din conul S de a genera o RTE sugerează că însumarea neuronală are loc în SC și este dependentă de aferențele vizuale transmise prin brahiul acestuia.
Există, astfel, dovezi convergente la pacienții cu creier divizat, la pacienții hemianopici cu orbire și din experimentele care utilizează stimuli cu lungime de undă scurtă, că însumarea neuronală are loc în SC. Cu toate acestea, în timp ce stimulii de lungime de undă scurtă nu activează celulele ganglionare retiniene care se proiectează către SC prin brahium, nu se poate concluziona că coliculul este în întregime orb la astfel de stimuli. Înregistrările unitare în SC la maimuțe au demonstrat că stimulii de lungime de undă scurtă activează răspunsurile în straturile superficiale ale coliculului. Mai mult decât atât, tractografia a demonstrat, la acei pacienți hemisferectomizați cu RTE (adică orbire), dar nu și la acei pacienți care nu prezentau semne de orbire, că SC avea conexiune cu emisfera intactă (Leh et al., 2006a).
Constatările investigației actuale oferă, astfel, prima dovadă directă că în SC are loc o însumare spațială care depinde de transmiterea semnalelor vizuale prin brahiul său.
Declarație etică
Acest studiu a fost realizat în conformitate cu orientările și cu aprobarea protocolului de către Comitetul de Etică al NHS, Bangor, Marea Britanie, și Comitetul de Etică al Școlii de Psihologie a Universității Bangor, Marea Britanie. Consimțământul informat oral și scris a fost obținut de la toți participanții, în conformitate cu Declarația de la Helsinki.
Contribuții ale autorilor
MvK: A conceput cercetarea, a proiectat și programat experimentul, a colectat și analizat datele pacienților și a contribuit la redactarea manuscrisului. KK: A colectat datele de control, a revizuit literatura relevantă, a analizat datele de control și a contribuit la redactarea manuscrisului. RR: A recrutat pacienții și a analizat studiile neuroimagistice, inclusiv disecția virtuală cu tractografia DTI, a pregătit istoricul pacienților și a contribuit la redactarea manuscrisului.
Declarație privind conflictul de interese
Autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
Behrens, T., Woolrich, M. W., Jenkinson, M., Johansen-Berg, H., Nunes, R. G., Clare, S., et al. (2003). Caracterizarea și propagarea incertitudinii în imagistica RM ponderată prin difuzie. Magn. Reson. Med. 50, 1077-1088. doi: 10.1002/mrm.10609
PubMed Abstract | Textul integral | Google Scholar
Behrens, T. E., Berg, H. J., Jbabdi, S., Rushworth, M. F., și Woolrich, M. W. (2007). Tractografia de difuzie probabilistică cu orientări multiple ale fibrelor: ce putem câștiga? Neuroimage 34, 144–155.
Google Scholar
de Gelder, B., Pourtois, G., van Raamsdonk, M., Vroomen, J., and Weiskrantz, L. (2001). Unseen stimuli modulate conscious visual experience: evidence from interhemispheric summation. Neuroreport 12, 385–391. doi: 10.1097/00001756-200102120-00040
CrossRef Full Text | Google Scholar
De Monasterio, F. M. (1978). Properties of ganglion cells with atypical receptive-field organization in retina of macaques. J. Neurophysiol. 41, 1435–1449.
PubMed Abstract | Google Scholar
Fries, W. (1984). Cortical projections to the superior colliculus in the macaque monkey: a retrograde study using horseradish peroxidase. J. Comp. Neurol. 230, 55–76. doi: 10.1002/cne.902300106
PubMed Abstract | CrossRef Full Text | Google Scholar
Georgy, L., Celeghin, A., Marzi, C. A., Tamietto, M., and Ptito, A. (2016). Coliculul superior este sensibil la configurația stimulilor de tip gestalt la pacienții cu hemisferectomie. Cortex 81, 151-161. doi: 10.1016/j.cortex.2016.04.018
PubMed Abstract | Full CrossRef Text | Google Scholar
Hershenson, M. (1962). Timpul de reacție ca măsură a facilitării intersenzoriale. J. Exp. Psychol. 63, 289-293. doi: 10.1037/h0039516
CrossRef Full Text | Google Scholar
Hikosaka, O., și Wurtz, R. (1983). Funcțiile vizuale și oculomotorii ale substantia nigra pars reticulata a maimuțelor. III. memorie contingentă a răspunsurilor vizuale și sacadate. J. Neurophysiol. 49, 1268-1284.
PubMed Abstract | Google Scholar
Leh, S. E., Johansen-Berg, H., și Ptito, A. (2006a). Viziunea inconștientă: noi perspective asupra corelației neuronale a vederii oarbe folosind tractografia de difuzie. Brain 129(Pt 7), 1822–1832.
PubMed Abstract | Google Scholar
Leh, S. E., Mullen, K. T., and Ptito, A. (2006b). Absence of S-cone input in human blindsight following hemispherectomy. Eur. J. Neurosci. 24, 2954–2960.
PubMed Abstract | Google Scholar
Marzi, C., Tassinari, G., Aglioti, S., and Lutzemberger, L. (1986). Spatial summation across the vertical meridian in hemianopics: a test of blindsight. Neuropsychologia 24, 749–758. doi: 10.1016/0028-3932(86)90074-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Miller, J. (1982). Divided attention: evidence for coactivation with redundant signals. Cogn. Psychol. 14, 247–279. doi: 10.1016/0010-0285(82)90010-X
CrossRef Full Text | Google Scholar
Miniussi, C., Girelli, M., and Marzi, C. A. (1998). Neural site of the redundant target effect electrophysiological evidence. J. Cogn. Neurosci. 10, 216–230. doi: 10.1162/089892998562663
CrossRef Full Text | Google Scholar
Pare, M., and Wurtz, R. (1997). Monkey posterior parietal cortex neurons antidromically activated from superior colliculus. J. Neurophysiol. 78, 3493–3497.
PubMed Abstract | Google Scholar
Poliva, O., Bestelmeyer, P. E., Hall, M., Bultitude, J. H., Koller, K., and Rafal, R. D. (2015). Cartografierea funcțională a cortexului auditiv uman: investigarea fMRI a unui pacient cu agnosie auditivă din cauza unei traume la coliculul inferior. Cogn. Behav. Neurol. 28, 160-180. doi: 10.1097/WNN.00000000000000000072
PubMed Abstract | CrossRef Full Text | Google Scholar
Posner, M., Snyder, C., and Davidson, B. (1980). Atenția și detectarea semnalelor. J. Exp. Psychol. Gen. 109, 160-174. doi: 10.1037/0096-3445.109.2.160
CrossRef Full Text | Google Scholar
Raab, D. H. (1962). Facilitarea statistică a timpilor de reacție simplă. Trans. N. Y. Acad. Sci. 24, 574-590. doi: 10.1111/j.2164-0947.1962.tb01433.x
CrossRef Full Text | Google Scholar
Reuter-Lorenz, P., Nozawa, G., Gazzaniga, M. S., și Hughes, H. C. (1995). Soarta țintelor neglijate: o analiză cronometrică a efectelor țintelor redundante în creierul bisectat. J. Exp. Psychol. Hum. Percept. Perform. 21, 211-230. doi: 10.1037/0096-1523.21.2.211
PubMed Abstract | CrossRef Full Text | Google Scholar
Savazzi, S. și Marzi, C. A. (2004). Coliculul superior subservește însumarea neuronală interhemisferică atât la persoanele normale, cât și la pacienții cu o secțiune totală sau agenezie a corpului calos. Neuropsychologia 42, 1608-1618. doi: 10.1016/j.neuropsychologia.2004.04.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Schiller, P. H., Malpeli, J. G., și Schein, S. J. (1979). Compoziția intrării geniculostriate ot coliculul superior al maimuței rhesus. J. Neurophysiol. 42, 1124-1133.
PubMed Abstract | Google Scholar
Sommer, M., și Wurtz, R. (2000). Compoziția și organizarea topografică a semnalelor trimise de la câmpul frontal al ochiului la coliculul superior. J. Neurophysiol. 83, 1979–2001.
PubMed Abstract | Google Scholar
Song, J. H., Rafal, R. D., and McPeek, R. M. (2011). PNAS Plus: deficits in reach target selection during inactivation of the midbrain superior colliculus. Proc. Natl. Acad. Sci. U.S.A. 108, E1433–E1440. doi: 10.1073/pnas.1109656108
PubMed Abstract | CrossRef Full Text
Tomaiuolo, F. (1997). Blindsight in hemispherectomized patients as revealed by spatial summation across the vertical meridian. Brain 120, 795–803. doi: 10.1093/brain/120.5.795
PubMed Abstract | CrossRef Full Text | Google Scholar
Zhaoping, L. (2016). From the optic tectum to the primary visual cortex: migration through evolution of the saliency map for exogenous attentional guidance. Curr. Opin. Neurobiol. 40, 94–102. doi: 10.1016/j.conb.2016.06.017
PubMed Abstract | CrossRef Full Text | Google Scholar