Există arbori genealogici și populații în care principiul împerecherii panmictice este în esență încălcat. De obicei, este vorba de comunități mici relativ izolate din punct de vedere geografic sau social. În aceste comunități, consangvinizarea, adică împerecherea consangvină de diferite ordine (un părinte – un copil, frați, verișori, unchi – nepoată etc.), poate fi larg răspândită.
Problema influenței consangvinității asupra populației este o problemă importantă a geneticii 1. Acum, această problemă nu a primit încă soluția finală.
În cele mai multe cazuri, consangvinizarea într-un arbore genealogic se ia în considerare pe trei generații. Cele mai îndepărtate rude sunt cele care sunt luate în considerare la împerecherea consangvină există verișori de gradul doi.
Consangvinizarea se ia în considerare cu ajutorul factorului de consangvinizare.
Factorul de consangvinizare este probabilitatea ca două alele să fie identice prin origine.
Să luăm în considerare metoda de calcul a factorului de consangvinizare într-un arbore genealogic oferită de Wright 1, 2.
Pentru aflarea factorului de consangvinizare este necesar:
să găsim pentru descendenții care se căsătoresc toate căile dintr-un arbore genealogic care prin strămoșul comun leagă acești descendenți.
să numărăm numărul de pași (alternanțe de generații) pe fiecare cale.
Apoi factorul de consangvinizare este egal cu:
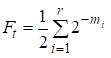 ……(1)
……(1)
unde r este numărul de căi, m1, m2, …. mi- numărul de pași pe fiecare cale.
Încă o dată vom observa că numărul de adaosuri din suma (1) este egal cu numărul de căi care prin strămoșii comuni se leagă căsătoria.
Să considerăm cazuri concrete de arbori genealogici, figura 1. În această figură pătratul semnifică un individ de sex masculin cercul – individ de sex feminin. Liniile punctate și cifrele identice precizează căile posibile într-un arbore genealogic pe care prin strămoșul comun (bărbatul sau femeia) se leagă de descendenți căsătoria consangvină. Pe unele site-uri de căi, figura 1с, există două cifre 1 și 2. Se specifică faptul că astfel de situri sunt cele comune pentru diferitele moduri.
Căsătoria un tată – fiică este incest, Figura 1а. Descendentul (femeia) este legat de strămoșul (bărbatul) acolo 1 cale cu un pas, adică m = 1. Prin urmare, există un factor de consangvinizare.
Ft = 1/2,2-1 = 1/4
Există căsătoria un frate – soră (sibs), Figura 1b. Cifrele precizează numărul de cale. Numărul de cifre identice pe drumul de la bărbatul care se căsătorește la femeie este egal cu numărul de pași.
Dacă fiecare descendent care s-a căsătorit se leagă cu partenerul pe două căi cu câte doi pași pe fiecare cale, adică m = 2. Prin urmare, există factor de consangvinizare.
Ft = 1/2.(2-2 + 2-2) = 1/4
Căsătoria între verișori este prezentată în Figura 1с.
Figura 1. Câteva tipuri de consangvinizare într-un arbore genealogic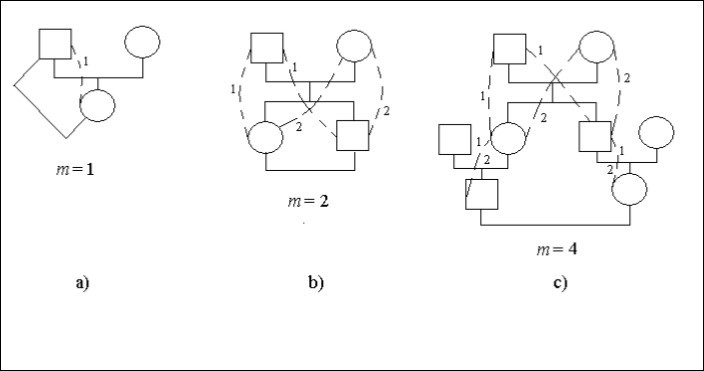
Care descendent care s-a căsătorit se leagă cu partenerul cele 2 căi cu 4 pași pe fiecare cale, adică m = 4. Prin urmare, există un factor de consangvinizare.
Ft = 1/2.(2-4 + 2-4) = 1/16
Formula (1) este ușor de înțeles din următoarea logică.
Evident, există 1/2 gene ale copilului este comună cu fiecare părinte, 1/4 – cu bunicul și bunica, 1/8 – cu străbunicul și străbunica, etc. Dacă de la femeie până la un strămoș oarecare duce drumul cu η pași partea de gene comune cu acest strămoș este egală cu 2-η. Dacă de la bărbat la orice strămoș conduce cu μ pași, cota de gene comune este egală cu 2-μ. La căsătoria bărbatului și a femeii cu strămoșul comun, ponderea genelor comune cu acest strămoș va fi egală cu 2-η. 2-μ = 2-m unde m = η + μ este numărul total de pași prin strămoșul comun de la femeie până la căsătoria bărbatului. Mărimea 1/2 2-m arată probabilitatea ca gena aleasă întâmplător la femeie să fie identică prin origine cu gena aleasă întâmplător la bărbat. Aceeași mărime indică contribuția la factorul de consangvinizare Ft un singur drum prin strămoșul comun de la femeie până la bărbat care se căsătorește.
De exemplu, dacă strămoșul comun va fi departe de căsătorie pe η = μ = 5 generații, probabilitatea ca ei să aibă gena comună este egală. 1/2 2-(5+5) = 1/2048 Această mărime la analiza împerecherii poate fi neglijată. (Tabelul 1)
Tabelul 1.factorii de consangvinizare pentru unele căsătorii consangvinizate sunt prezentați în plus.
| Tipul de consangvinitate căsătorie | Factorul de consangvinizare Ft |
| Prieteni de gradul doi | 1/32 |
| Unchiul văr – niece | 1/32 |
| Third cousins | 1/64 |
The inbreeding factor Ft is the probability of two alleles in posterity are identical by origin i.e. occur from same gene of ancestor 1. Or, that too most, the inbreeding factor is a probability of the allele origin in posterity is consanguineous. Hence, the probability of the non-consanguineous origins of identical alleles in homologous chromosomes is equal 1 – Ft.
Let’s find total probability of the recessive homozygote aa occurrence (eventаа) at random mating for an autosomal genes 3:
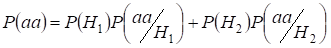 …….(2)
…….(2)
This formula follows from the following logic of mating: originea alelelor identice în autosomi omologi este neconsangvină (probabilitatea acestei ipoteze este P(H1) = 1- Ft )apare și homozigotаа (probabilitatea condiționată a acestui eveniment este P(aa/H1) = q2) sau originea alelelor identice este consangvină (probabilitatea acestei ipoteze este P(H2) = Ft ) și apare homozigotааа (probabilitatea condiționată a acestui eveniment este P(aa/H2) = q). Probabilitatea q de apariție a homozigotului aa în acest din urmă caz este determinată de faptul că dacă alela a se află într-un autosom, atunci la originea consangvină a homozigotului aceeași alelă trebuie să fie în mod necesar și în autosomul omolog.
În consecință, probabilitatea totală de apariție a homozigotului recesiv este egală cu:
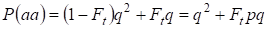 ……….(3)
……….(3)
unde se ia în considerare p = 1 – q există frecvența alelei dominante a.
În mod similar se găsește probabilitatea totală de apariție a homozigotului dominant AA la împerechere aleatorie:
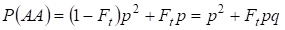 ……….. (4)
……….. (4)
Prin urmare, conform (3) și (4) frecvența totală a homozigoților (dominanți și recesivi) în arborele genealogic consangvinizat crește pe 2Ftpq. Prin urmare, frecvența heterozigoților scade pentru aceeași dimensiune și pentru a deveni egală cu 2pq – 2pqFt = 2pq(1-Ft), deoarece suma frecvențelor tuturor zigoților este egală cu 1.
Având în vedere factorul de consangvinizare și, de asemenea, (3) și (4), legea Hardy – Weinberg în următoarea generație a arborelui genealogic consangvinizat pentru un genom autosomal trebuie să se scrie distribuțiile genotipurilor zigoților ca 4:
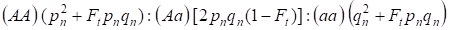 …………(5)
…………(5)
Unde pn este frecvența alelelor dominante A în generația n, qn – frecvența alelelor recesive a, deci pn + qn = 1.
Raportul (5) se referă și la raportul lui Wright 5. Raportul lui Wright arată că într-un arbore genealogic consangvin în urma împerecherii consangvinizate a indivizilor se observă următoarele efecte: se constată creșterea fracției homozigoților autosomali într-un arbore genealogic pe Ft pnqnși reducerea fracției heterozigoților autosomali în cele (1-F1) în comparație cu o populație panmictică.
Dar deja printr-o generație se restabilește echilibrul Hardy – Weinberg care specifică importanța mică a împerecherii consangvinice separate pentru un arbore genealogic:
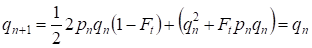 ………(6)
………(6)
Evident, pentru o populație în ansamblu, actul separat al împerecherii consangvinice va avea o importanță și mai mică decât pentru un arbore genealogic.
Același efect de variație a frecvenței genotipurilor zigoților se observă și pentru genele legate de sex.
Distribuția genotipurilor legate de cromozomul Х în arborele genealogic panmic este descrisă de legea Hardy – Weinberg ca fiind 1:
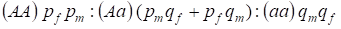 ………..(7)
………..(7)
Frecvența unei alele dominante A o vom desemna la bărbați pm și la femei pf. Pentru alelele recesivea este în mod corespunzător qm și qf .
La împerechere există un raport al genotipurilor la femei conform produsului (pf + qf) (pm +qm).
Să găsim probabilitatea totală de apariție a homozigotului recesiv aa la împerecherea aleatorie pentru genele fiicei legate de cromozomulХ conform formulei (2). Logica evenimentelor este similară cu cazul autosomelor, cu excepția faptului că probabilitatea condiționată de apariție a unui genotipаа de la alele neconsanguine conform (7) este egală cu P(aa/H1) = qmnqfn. Pentru alelele consangvine, probabilitatea condiționată a apariției unui genotipаа este egală cu P(aa/H2) = qf(n+1). La fiice, adică în generația n+1, probabilitatea de apariție a homozigotului aa este determinată de faptul că, dacă alela a se află într-un cromozom Х, atunci la originea consangvină a homozigotului aceeași alelă trebuie să fie în mod necesar și în cromozomul Х omolog.
În consecință, probabilitatea totală de apariție a homozigotului recesiv este egală:
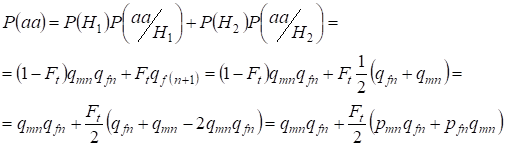 ……(8)
……(8)
unde pmn este frecvența alelelor dominante A în generația n la bărbați și pfn – la femei. Pentru alelele recesive a în consecință qmn și qfn . În plus se folosește qmn =1 – qmn și pfn = 1 – qfn.
În mod similar găsirea probabilității totale de apariție a homozigoților dominanți cu utilizarea:
 ,
,
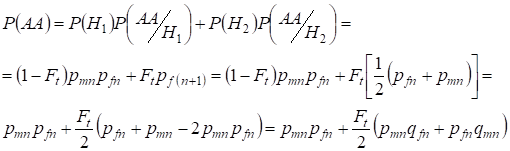 ……….(9)
……….(9)
și un heterozigot P(Aa) = 1- P(aa) – P(AA) vom găsi distribuția genotipurilor în cromozomii Х din arborele genealogic consangvinizat la împerechere:
 ……..(10)
……..(10)
Se demonstrează simultan că, la fel ca și în autosomi, aditivii consangvini în frecvențele homozigoților dominanți și recesivi în cromozomii legați de sex sunt identici.
O altă modalitate de constatare a raportului (10) va consta în cele ce urmează.
Să găsim mai întâi probabilitatea totală de apariție a unui heterozigot Aa, adică P(Aa) cu ocazia împerecherii consangvinice aleatoare. Această probabilitate rezultă din următoarea logică a împerecherii: originea alelelor este neconsangvină (probabilitatea acestei ipoteze este P(H1) = 1 – Ft ) apare și heterozigotul Aа (probabilitatea condiționată a acestui eveniment este P(Aa/H1) = pmnqfn + pfnqmn, a se vedea formula (7)) sau originea alelelor este consangvină (probabilitatea acestei ipoteze este P(H2) = Ft) și apare heterozigotul Aа (probabilitatea condiționată a acestui eveniment este egală cu zero P(Aa/H2) = 0 ). Ultima afirmație se definește prin aceea că la origine consangvină nu pot apărea alele similare în cromozomi Х omologi nu poate apărea heterozigotulАа format din alele diferite.
De aceea, folosind formula probabilității totale găsim probabilitatea totală de apariție a heterozigotului Aa:
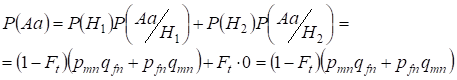 ……..(11)
……..(11)
Reducerea frecvenței heterozigoților Aa are loc, așa cum s-a demonstrat anterior, datorită creșterii identice a frecvenței homozigoților dominant AA și recesiv aa. De exemplu, creșterea frecvenței homozigoților aa recesivi este egală:
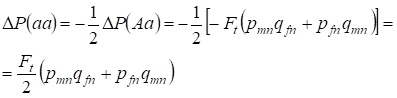 ……..(12)
……..(12)
unde reducerea frecvenței heterozigoților Aa în raport cu arborele genealogic panmic conform (7) și (11) este egală. ΔP(Aa) = – Ft (pmnqfn + pfnqmn) Utilizând (11) și (12) pentru a actualiza (7) pentru cazurile de consangvinizare, găsim distribuția genotipurilor (10) în cromozomii Х în arborele genealogic consangvinizat al fiicelor.
În generația următoare frecvența alelelor recesive a la femei este egală:
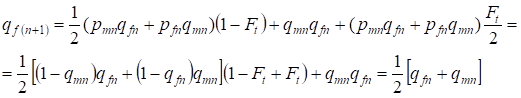 ……..(13)
……..(13)
Ceea ce respectă întru totul aceeași frecvență la absența consangvinității, adică se restabilește echilibrul Hardy – Weinberg sau este mai exact apropierea de echilibru.
Raportul (7) și (13) arată în ansamblu că frecvențele alelelor într-un arbore genealogic pentru contul de consangvinizare nu variază. Există doar o redistribuire a alelelor. O parte din alele părăsesc heterozigoții și trec la homozigoți.
.