1. Řasinky
2. Bičíková tělíska
3. Struktura
4. Pohyb
5. Vznik
Mikrotubuly jsou součásti cytoskeletu s důležitými funkcemi ve fyziologii buňky. Mikrotubulární lešení cytoplazmy je díky polymerizační a depolymerační schopnosti mikrotubulů velmi plastické. Ne všechny buněčné mikrotubuly však podléhají tomuto stádiu smršťování nebo růstu. Cilie, bičíky a centrioly/základní tělíska jsou buněčné struktury obsahující velmi stabilní (počtem i délkou) a vysoce organizované mikrotubuly. Na této stránce se budeme zabývat řasinkami a bičíky.
1. Řasinky a bičíky. Řasinky
Řasinky jsou tenké a dlouhé buněčné výběžky o průměru asi 0,25 µm a délce asi 10 až 15 µm, které se vyskytují u živočišných buněk a některých jednobuněčných eukaryotických druhů. Obvykle jsou těsně zabaleny na volném povrchu epitelových buněk (obr. 1 an 2), jako je epitel dýchacích cest, epitel rozmnožovacích kanálků, žábry ryb a mlžů atd. Řasinky jsou pohyblivé struktury a jejich hlavní funkcí je pohybovat okolní tekutinou, jako je hlen povrchu dýchacích cest, voda kolem epitelu žaber, ale také oocyt v samičím vejcovodu. Mnoho jednobuněčných organismů se může pohybovat poháněno řasinkami a jiné je mohou využívat k vytváření vodního víru pro zachycení potravy. Uzlové řasinky embrya se podílejí na iniciaci levopravé osy během embryonálního vývoje obratlovců. Pohyb řasinek je podobný tlukotu, při kterém dochází k impulsu kapaliny rovnoběžně s povrchem buňky.
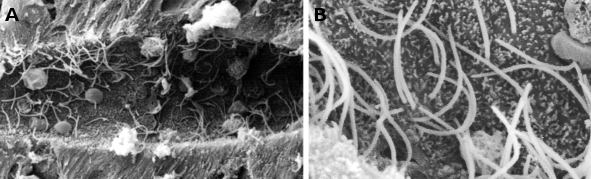
Obr. 1. Snímky ze skenovací elektronové mikroskopie zobrazující centrální kanál míchy mihule. Lze pozorovat mnoho řasinek (při větším zvětšení v B) a drobné mikrovily v apikální oblasti ependimálních buněk.
Obrázek 2. Snímky respiračního epitelu z transmisní elektronové mikroskopie. Buňky s průhlednou cytoplazmou vykazují na svém apikálním povrchu mnoho řasinek.
Existují řasinky, které se nemohou pohybovat, a proto nejsou určeny k pohybu tekutin. Tyto řasinky se označují jako primární řasinky. Většina dosud studovaných buněk (s výjimkou červených krvinek) nese primární řasinky: buňky vaječníku, neurony, chondrocyty, buňky ektodermu, mezenchymální buňky, epitelové buňky močových cest, hepatocyty a dokonce i kultivované buňky. Původně byly primární řasinky považovány za nefunkční řasinky. Na řasinkové membráně však bylo nalezeno mnoho typů receptorů a iontových kanálů, takže byly považovány za buněčné senzorické struktury. Například čichové receptory se nacházejí v řasinkách jejich dendritů a vnější segmenty tyčinek a čípků sítnice jsou vlastně modifikované řasinky. Některé receptory jsou v řasinkové membráně více zabaleny než v jiných doménách plazmatické membrány. Kromě toho se v nitru řasinek nachází široká škála molekul, které se podílejí na rolích přenosu signálu. Díky vyššímu poměru povrch/objem řasinky jsou intraciliární molekulární reakce intenzivnější a účinnější, než kdyby byly vně řasinky. Kromě chemického signálu mohou primární řasinky detekovat pohyb tekutiny mimo buňku a fungovat jako mechanoreceptory.
2. Vlivem chemického signálu se řasinky mohou pohybovat i mimo buňku. Bičíky
Bičíky jsou podobné řasinkám, ale jsou mnohem delší, asi 150 µm dlouhé, a o něco silnější. V buňkách jsou poměrně méně početné než řasinky. Hlavní funkcí bičíků je pohyb buňky. Pohyb bičíků se liší od pohybu řasinek, protože směr pohybu je kolmý k povrchu buňky (nikoliv rovnoběžný), tedy směr podélné osy bičíku. Bičíky lze často pozorovat u pohyblivých buněk, jako jsou jednobuněčné organismy a spermie.
Struktura
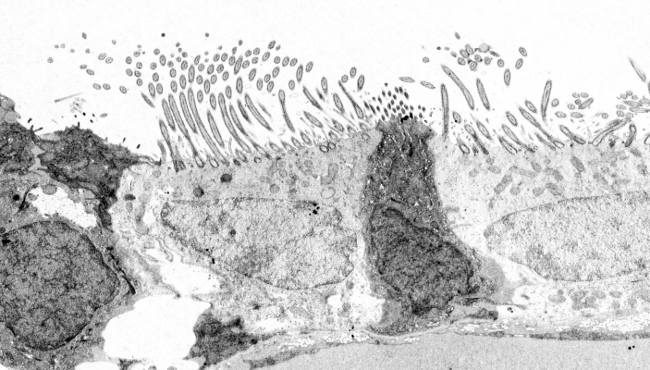
Cílie a bičíky jsou složité struktury obsahující více než 250 různých proteinů. Obě mají stejnou centrální organizaci mikrotubulů a další přidružené proteiny, dohromady známé jako axonemy, a jsou ohraničeny plazmatickou membránou (obr. 3). Kromě axonemu se uvnitř řasinek/bičíků nachází mnoho rozpustných molekul tvořících matrix. Axoném se skládá z 9 párů mikrotubulů kolem dalšího centrálního páru mikrotubulů. Toto uspořádání lze zapsat jako (9 x 2) + 2. Primární řasinky postrádají centrální pár mikrotubulů. Každý mikrotubul centrálního páru je tvořen 13 protofilamenty, ale mikrotubuly periferních párů mezi sebou sdílejí některá protofilamenta. Periferní pár je tedy tvořen mikrotubuly A a B. Mikrotubulus A obsahuje 13 protofilament a mikrotubulus B obsahuje 10 nebo 11 protofilament, přičemž 2 nebo 3 sdílí s mikrotubulem A.

Obrázek 3. Hlavní molekulární složky řasinek a bičíků. U primárních řasinek centrální pár chybí
Mikrotubulární organizace axonemu je výsledkem lešení z proteinů. Jako složky axonemu, které se podílejí na udržování mikrotubulární organizace, bylo nalezeno již dvanáct proteinů. Sousední periferní páry mikrotubulů jsou mezi sebou spojeny nexinem. V každém páru je mikrotubul A spojen proteinovými paprsky s centrálním prstencem, který obsahuje centrální pár nebo mikrotubuly. Dinein je motorický protein spojený s periferními mikrotubuly, který se podílí na pohybu řasinek a bičíků.

Obrázek 4. Řasinky a bičíky. Ultrastruktura řasinky ependimální buňky míchy. (9+2)x2 znamená 9 periferních párů a 1 centrální pár mikrotubulů.
Mikrotubuly jsou polymerovány z bazálních tělísek (obrázky 3 a 4). Bazální tělísko se skládá z 9 tripletových mikrotubulů tvořících válec (podobně jako centrioly). Chybí jim centrální pár mikrotubulů, takže je to (9×3)+0. V každém tripletu má pouze jeden mikrotubul (mikrotubul A) kompletní sadu protofilament, zatímco mikrotubuly B a C jich mezi sebou část sdílejí. Z bazálního tělíska vyrůstají mikrotubuly A a B a tvoří periferní mikrotubuly axonemu. Těsně nad bazálním tělískem se nachází oblast řasinek známá jako přechodová zóna, která obsahuje 9 periferních párů a žádný centrální pár. Bezprostředně za přechodovou zónou se nachází bazální destička, ze které polymeruje centrální pár mikrotubulů, čímž je axoném dokončen. Všechny mikrotubuly mají plusový konec směrem ke špičce řasinky/bičíku. Proximální konec bazálního tělíska (vnitřní neboli minusový konec mikrotubulů) je ukotven k buněčnému cytoskeletu prostřednictvím dlouhých bílkovinných vláken nazývaných řasinkové kořínky
Kromě axonému mají řasinky/flagella další oddíly. Membrána obsahuje mnoho receptorů a kanálků pro vnímání prostředí, speciálně u primárních řasinek. Tekutá fáze vnitřku se nazývá matrix, která pomáhá s udržováním organizovanosti celé struktury a je zodpovědná za přenos informací získaných membránovými receptory. Dalšími odlišnými oblastmi jsou bazální tělísko nacházející se u základny a apikální část řasinky/flagellum, která obsahuje proteiny stabilizující plusové konce mikrobuněk.
4. Řasinka, která se nachází na bázi řasinky. Pohyb
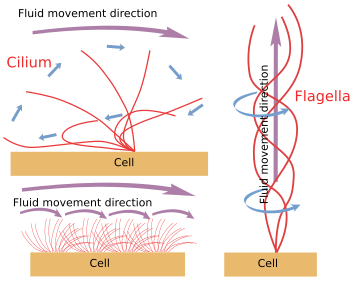
Obrázek 6. Cilium a flagela. Modely pohybu cilií a bičíků. Vytvářejí různé směry pohybu tekutin.
Když se cilium/flagella mechanicky oddělí od buňky, pokračují v pohybu, dokud se nevyčerpá zásoba ATP. To znamená, že mechanismus pohybu je vnitřní (obr. 6). Ve skutečnosti je pohyb vyvolán posunutím periferního páru mikrotubulů po sousedním. Proteiny Nexin a spoke zabraňují dezorganizaci axonemu, ale umožňují tyto pohyby. Dinein je motorický protein zodpovědný za posuvný pohyb. Energii mu dodává ATP. Dineiny jsou ukotveny svou globulární částí k mikrotubulu A jednoho periferního páru a svou ocasní částí k mikrotubulu B sousedního páru. Molekulární mechanismus je podobný jako u cytosolových dineinů, ale místo transportu nákladu se pohybuje mikrotubuly. Pro efektivní pohyb je nutná koordinace dineinů axonemu. Vlny vápníku v nitru řasinky/bičíku mohou koordinovat aktivaci dineinů a v případě potřeby měnit frekvenci pohybu. Je třeba si uvědomit, že ne všechny dineiny musí být aktivovány současně, ale synchronně.
5. Způsoby aktivace dineinů Tvorba
Během diferenciace si buňky vytvářejí všechny řasinky a bičíky potřebné pro jejich normální fyziologii. To znamená, že všechny musí být vytvořeny od základu. Mikrotubuly axonémů jsou nukleovány z mikrotubulů A a B bazálních tělísek, takže na jedno řasnaté/bičíkové tělísko je potřeba jedno bazální tělísko. Jak vzniká více bazálních tělísek? Existují nejméně tři způsoby tvorby bazálních tělísek: a) pomocí centriol jako šablon pro nukleaci bazálních tělísek; b) z amorfního materiálu známého jako deuterozom; c) u rostlin existují odlišné bílkovinné agregáty, které mohou nukleovat bazální tělíska.
Důsledkem vad řasinek/flagel jsou četné lidské patologie. Označují se jako ciliopatie a patří mezi ně náhodná lateralita, nesprávné uzavření neurální trubice, polydaktylie, cystické patologie ledvin, jater a slinivky břišní, degenerace sítnice, obezita a kognitivní vady.
Bibliografie
Marshall WF, Nonaka S. 2006. Cilie: naladění buněčné antény. Současná biologie. 16:R604-R614.
Satir P, Christensen ST. 2007. Přehled struktury a funkce řasinek savců. Annual review of phisiology. 69:377-400.