Úvod
Dva zrakové signály, které se objeví současně, jsou detekovány rychleji než kterýkoli z nich (Hershenson, 1962; Raab, 1962). Část tohoto efektu redundantního cíle (RTE) lze stochasticky vysvětlit pomocí modelu „koňských dostihů“. To znamená, že pokud jsou dva podněty zpracovávány v paralelních a nezávislých kanálech, u nichž se rychlost zpracování náhodně mění od pokusu k pokusu, při každé prezentaci vyhrává závod nejrychlejší podnět, který dosáhne prahu detekce a spustí reakci; proto je v průměru pravděpodobnější, že dva podněty přinesou rychlejší reakci než průměrná doba reakce na jeden podnět zpracovávaný v jednom z obou kanálů.
Analýzy kumulativních frekvenčních distribucí však ukázaly, že existuje další příspěvek k RTE, který nelze zohlednit modelem dostihu mezi dvěma nezávislými kanály. Miller (1982) navrhl „koaktivační“ model, v němž se oba signály sčítají v aktivačním fondu. Miniussi et al (1998) ukázali, že redundantní cíle způsobují kratší latence pro mozkové potenciály související s událostmi P1 a N1, což naznačuje, že k nervové sumaci dochází na počátku zrakové dráhy. Experimenty u hemianopických pacientů také ukázaly, že RTE nelze plně vysvětlit závodem o dosažení prahu detekce, protože RTE se může projevit i při absenci fenomenálního uvědomění si jednoho ze dvou podnětů (Marzi et al., 1986; Tomaiuolo, 1997; de Gelder et al., 2001; Leh et al., 2006a,b). U pacientů po hemisférektomii totiž redundantní cíl ve slepém poli nejenže generuje RTE, ale RTE se zvyšuje, když redundantní podnět doplňuje gestaltový vzorec (Georgy et al., 2016). Tato pozorování u hemianopiků naznačují, že nervová sumace probíhá subkortikálně a není závislá na primární zrakové kůře.
Kumulativní nepřímé důkazy implikují superior colliculus (SC) jako substrát pro nervovou sumaci, která přispívá k RTE. Přímý důkaz však dosud nebyl podán. Současný výzkum zkoumá nervovou dráhu, která přenáší zrakové signály, jež jsou integrovány v SC. SC se skládá z povrchových a hlubokých vrstev. Hluboké vrstvy koliculu přijímají aferenty, a to jak přímé, tak prostřednictvím bazálních ganglií, z okulomotorické kůry (Hikosaka a Wurtz, 1983; Pare a Wurtz, 1997; Sommer a Wurtz, 2000). Zrakové aferenty do povrchových vrstev koliculu jsou však přenášeny jak ze sítnice (prostřednictvím retinotektálního traktu), tak z časné zrakové kůry (Fries, 1984), a to přes brachium SC.
Protože u neurologických pacientů, kterým chybí zraková kůra, byly prokázány efekty nervové sumace, mohly by se zrakové signály sumované v koliculu přenášet přímo ze sítnice do povrchových vrstev koliculu prostřednictvím retinotektálního traktu. Pokud tomu tak je, mohla by být nervová sumace zrušena lézí raménka SC.
Léze, které narušují raménko SC, jsou velmi vzácné. Zde uvádíme jednotlivé případy dvou pacientů s jednostrannými subkortikálními lézemi, které deaferentovaly SC od zrakového vstupu a u kterých se neprojevila RTE. Jeden pacient měl lézi poškozující jak pulvinární jádro talamu, tak rostrální SC a jeho brachium. U druhého pacienta s poškozením dorzálního středního mozku v důsledku traumatického poranění mozku se SC jevil jako neporušený, ale jeho raménko bylo narušeno.
Materiál a metody
Účastníci
Dva neurologičtí pacienti, každý s vlastní kontrolní skupinou odpovídající svému věku, byli testováni v experimentu k měření RTE.
RE bylo v době testování 71 let. Zadní krvácení do thalamu před 3 lety zničilo mediální pulvinární, ventrolaterální thalamická jádra a zadní končetinu vnitřní kapsuly s poškozením zasahujícím do dorzálního středního mozku včetně pretekta a rostrálního SC (obr. 1) Pravděpodobnostní DTI traktografie (Behrens et al., 2003, Behrens et al., 2007) potvrdila, že léze zničila brachium SC (obr. 2). RE byla ochrnutá na levou stranu těla se ztrátou citlivosti v levém obličeji a paži. Měla abnormální pohyby očí včetně zatahování víček, paralyzované vertikální pohyby očí a poruchu konvergence, které byly důsledkem rozšíření léze do dorzálního středního mozku.
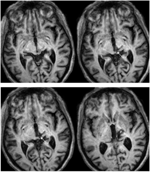
OBRÁZEK 1. T1 vážená MRI s vysokým rozlišením (0,7 mm3) pacientky RE. Axiální řezy od ventrálního (vlevo nahoře) po dorzální (vpravo dole) zobrazující lézi v pravém dorzálním středním mozku a thalamu. Rozšíření léze do rostrálního horního koliculu (SC) znázorňují bílé šipky.
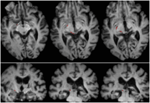
Obrázek 2. Traktografie zobrazující retinotektální trakt u pacienta RE koregovaný s T1 váženými axiálními řezy (nahoře od ventrálního k dorzálnímu) a koronálními řezy (dole od předního k zadnímu). Průběh retinotektálního traktu je zobrazen v levé hemisféře (červeně). V pravé hemisféře nebyla sledována žádná proudnice a při srovnání s intaktní hemisférou je vidět, že léze zničila brachium SC v pravé hemisféře. Červeně zobrazená proudnice byla vytvořena pomocí pravděpodobnostní traktografie s využitím FSL FDT (FMRIB Diffusion Toolbox; Behrens et al., 2003, Behrens et al., 2007; http://fsl.fmrib.ox.ac.uk/fsl/fslwiki). Difuzně vážené echo-planární snímky magnetické rezonance byly pořízeny v rozlišení 1,5 mm3 s 32 izotropně rozloženými směry kódování difuze (b = 800) a základní linií (b = 0). Doba opakování = 2 s a doba echa = 35 ms. Pro pravděpodobnostní traktografii jsme na optickém traktu každé hemisféry těsně za chiasmem ručně označili výchozí oblast (tj. nakreslili seed masky) a na SC cílovou oblast (target mask).
ML bylo v době testování 23 let. Před 7 lety utrpěla těžké traumatické poranění mozku při dopravní nehodě, které mělo za následek difuzní axonální poranění s krvácením do pravého putamen a dorzálního středního mozku. Podrobnosti o její anamnéze, neurologickém vyšetření a neurozobrazovacích nálezech uvádí (Poliva et al., 2015). Ačkoli vlevo byla určitá motorická porucha, včetně spasticity a ataxie, znovu získala nezávislost. Jejím hlavním postižením byla těžká sluchová agnozie v důsledku poškození sluchových drah mozkového kmene včetně levého dolního koliculu. Zraková ostrost a zorné pole byly neporušené a nedošlo k vizuálnímu zanedbání nebo vymizení. Okulomotorické příznaky dysfunkce středního mozku zahrnovaly makrovlnné záškuby a konvergenční křeč při vertikálním pohledu (více dolů než nahoru.) Nebyla zjištěna ptóza ani zornicová abnormalita. Magnetická rezonance s vysokým rozlišením prokázala cystickou dutinu v pravém putamen v místě jejího předchozího krvácení a malou periventrikulární lézi na pravém dolním ponsu v oblasti dolního mozečkového pedunklu (obr. 3). Měla téměř úplnou avulzi levého dolního koliculu, ušetřena byla pouze jeho nejmediálnější a kaudální část. Poškození zasahovalo do laterálního pretekta a tegmenta středního mozku a ničilo brachii horního a dolního koliculu a ventrálně do červeného jádra. Pravděpodobnostní traktografie DTI potvrdila, že léze zničila brachium SC (obr. 4).
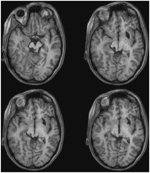
Obr. 3. MRI s vysokým rozlišením (0,7 mm3) T1-vážené zobrazení pacienta ML. Axiální řezy od ventrálního (vlevo nahoře) po dorzální (vpravo dole) zobrazující poškození levého dolního koliculu a mezencefalického tegmenta ventrálně a laterálně od SC.
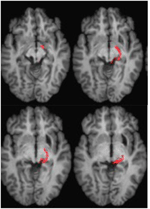
FIGURE 4. DTI traktografie u pacienta ML zobrazující retinotektální trakt v pravé hemisféře (červeně) koregistrovaný s T1 váženými MR obrazy v axiální rovině od ventrální (vlevo nahoře) po dorzální (vpravo dole). Difuzně vážené echo-planární obrazy magnetické rezonance byly pořízeny s rozlišením 2 mm3 s 63 izotropně rozloženými směry kódování difuze (b = 1000) a základní linií (b = 0). Doba opakování = 2 s a doba echa = 35 ms. Traktografie byla provedena podle popisu na obrázku 2.
Neurologicky zdravý kontrolní subjekt pro pacienta RE zahrnoval 11 starších dospělých (šest žen a pět mužů, průměrný věk: 68,5 let, rozmezí 59-73 let). Byli rekrutováni z panelu komunitních účastníků Bangorské univerzity. Účastníci neměli žádné známé neurologické, psychologické, psychiatrické nebo kognitivní poruchy a všichni účastníci měli normální/korigovaný až normální zrak. Účastníci obdrželi za svou účast platbu ve výši 6 liber.
Neurologicky zdravými kontrolními subjekty pro pacienta ML bylo 11 studentů a doktorandů Bangorské univerzity ve věku od 18 do 26 let (sedm žen a čtyři muži).
Aparatura a podmínky zobrazení
Programovací software Presentation® spuštěný na počítači PC zaznamenával reakční čas (RT) reakcí a generoval podněty, které byly prezentovány na monitoru Dell (12,5″ × 25″, obnovovací frekvence 60 Hz.) umístěném ve výšce očí 57 cm před účastníky s binokulárním zobrazením v podmínkách tlumeného osvětlení. Pro zajištění stability hlavy byla použita opěrka brady a hlavy. Reakce účastníků byly zaznamenávány pomocí stisknutí mezerníku na klávesnici.
Stimuly
Experimentální stimuly se skládaly z nevyplněných bílých značkovacích čtverců (1 cm × 1 cm) umístěných 8° vlevo a vpravo od malého fixačního pole (0,2 cm × 0,2 cm) ve středu obrazovky. Cílové podněty spočívaly ve vyplnění jednoho nebo obou okrajových značkovacích políček tak, aby vznikl celistvý bílý čtverec. Všechny podněty byly bílé a byly prezentovány na šedém pozadí.
Obrázek 5 ukazuje pořadí jednoho pokusu. Značková pole zůstala viditelná po celou dobu experimentu. Po intervalu mezi pokusy v délce 1750 ms se objevil fixační rámeček, který zahájil pokus. Po náhodném intervalu v rozmezí 250 až 750 ms (v krocích po 25 ms) se v místě jednoho nebo obou značkovacích políček objevil cíl vyžadující jednoduchou reakci stisknutím klávesy mezerníku na klávesnici. Cílem byl vyplněný bílý čtverec vytvořený vyplněním jednoho nebo obou políček značky. Náhodně a se stejnou pravděpodobností se cíle objevovaly vlevo, vpravo nebo na obou místech současně. Při pokusech s chytáním (10 % pokusů) se neobjevil žádný cíl.
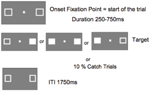
OBRÁZEK 5. Při pokusech s chytáním se neobjevil žádný cíl. Sekvence zobrazení jednoho pokusu (ITI, mezipokusový interval mezi reakcí účastníka a nástupem fixačního bodu, kterým začíná další pokus).
Postup
Každý účastník byl testován v jednom sezení. Po přečtení pokynů na obrazovce byly účastníkům ukázány příklady všech podnětů, které byly v úloze prezentovány (fixační pole a každá ze tří prezentací cílů), a byli požádáni, aby potvrdili rozpoznání každého cíle stiskem klávesy mezerník. Instrukce zněly, aby po dobu trvání každého pokusu udržovali fixaci na středovém rámečku a aby co nejrychleji reagovali stisknutím mezerníku na klávesnici pravou rukou, jakmile se objeví cíl.
Poté, co účastník ústně a prostřednictvím stisknutí klávesy potvrdil, že všem instrukcím porozuměl, pokračovalo se prezentací cvičných pokusů (10 pokusů z každé podmínky prezentace cíle a 3 pokusy s absencí cíle „catch“). Během dokončování cvičných pokusů byla na obrazovce prezentována zpětná vazba obsahující následující výroky: (1) „Správně“, (2) „Snažte se reagovat rychleji!“ a (3) „Reagujte pouze v případě, že vidíte cíl“. Jakmile bylo cvičení dokončeno a účastník i experimentátor si byli jisti, že instrukce k úkolu byly správně pochopeny, byl účastník vyzván k zahájení experimentu. Prvních 30 pokusů každého experimentálního bloku bylo vyloučeno jako cvičné pokusy. Každý blok se skládal z celkem 233 pokusů: 70 pokusů vpravo, 70 pokusů vlevo, 70 pokusů v obou směrech a 23 „záchytných“ pokusů. V polovině bloku byla udělena krátká přestávka. Pacient ML a mladší kontrolní skupina byli testováni v jednom bloku. Vzhledem k variabilitě související s věkem a stavem pacientky RE byla testována ve dvou blocích v různých dnech; a její věkově odpovídající kontrolní účastníci byli rovněž testováni ve dvou sezeních v jeden den.
Analýzy
Pro každou kontrolní skupinu byl vypočten medián RT pro každého účastníka v každém stavu (levý cíl, pravý cíl, oboustranné cíle) po vyloučení pokusů následujících po záchytných pokusech a pokusů s RT <100 ms nebo >800 ms. Byl proveden párový výběrový t-test, aby se potvrdilo, že neexistuje asymetrie pro odpovědi na pravé a levé jednostranné cíle, a byl vypočten průměrný RT pro jednostranné cíle. RTE byl vypočten odečtením RT pro bilaterální pokusy od RT pro unilaterální pokusy. Párový výběrový t-test porovnal unilaterální a bilaterální reakce, aby se zjistilo, zda je RTE statisticky spolehlivý.
U každého pacienta se po vyloučení pokusů následujících po záchytných pokusech a pokusů s RT <100 ms nebo >800 ms porovnal párový výběrový t-test medián RT pro kontralaterální a ipsilaterální unilaterální cíle. Jak je uvedeno níže, RT pro odpovědi na kontralaterální cíle byly delší než pro ipsilaterální unilaterální cíle. Ke zjištění, zda přítomnost kontralezionálního cíle vyvolala statisticky spolehlivý RTE, byly párovým výběrovým t-testem porovnány mediány RT pro pokusy s bilaterálním cílem s pokusy s unilaterálním ipsilaterálním cílem.
Pro ověření, zda byl RTE u každého pacienta spolehlivě menší než u jeho příslušné kontrolní skupiny, byl pro každého účastníka kontroly vypočítán RTE a vypočteny horní a dolní hranice 95% intervalů spolehlivosti (CI). Z-skóre pro RTE každého pacienta bylo vypočteno vzhledem k jeho kontrolní skupině a testováno na statistickou spolehlivost pomocí Crawfordova t-testu.
P hodnoty pro párové výběrové t-testy byly uvedeny pro dva chvosty a pro Crawfordův t, jeden chvost.
Výsledky
Pacienti RE
Kontrolní účastníci: Průměrné mediány RT se nelišily pro levé a pravé odpovědi (t = 0,6.) Obrázek 6 ukazuje, že RT byly kratší (RTE) pro bilaterální cíle než pro levé (t = 6,8, p < 0,001, nebo pravé (t1,10) = 3,8, p = 0,003) unilaterální cíle. Medián RT pro pokusy s bilaterálními cíli byl odečten od průměru mediánu RT pro unilaterální cíle, aby se vypočítal průměrný RTE pro každého účastníka kontroly. Průměrný RTE kontrolní skupiny byl 24,3 ms (SEM = 2,75).

Obr. 6. Medián RT u pacientů RE a průměr mediánu RT u účastníků kontrolní skupiny pro cíle objevující se v levém zorném poli („Left“), v pravém zorném poli („Right“) a současně v obou zorných polích („Both“). Chybové úsečky u účastníků kontrolní skupiny označují standardní chyby.
Pacient RE měl medián RT delší než kontrolní skupina (obr. 6) a byl nad horní hranicí 95% CI průměrných RT kontrolní skupiny pro všechny tři podmínky (levá = 457 ms; pravá = 449 ms; obě = 426 ms). RT na jednostranné cíle v kontralezionálním (levém) poli byly delší než u jednostranných cílů v ipsilezionálním (pravém) poli (t = 6,0, p < 0,001).
Důležité je, že nedošlo k RTE: Medián RT pro bilaterální cíle (500 ms) nebyl kratší než pro unilaterální cíle v ipsilesionálním (pravém) zorném poli (494 ms; t = 1,141), p = ns (obrázek 6). Tabulka 1 (nahoře) ukazuje, že RTE vypočtená pro pacientku RE byla významně nižší než RTE pro její kontrolní skupinu.

TABULKA 1. RTE vypočtená pro pacientku RE byla významně nižší než RTE pro její kontrolní skupinu. Zbytečný cílový efekt pro pacienty RE (nahoře) a ML (dole) a průměrné RTE pro jejich příslušné kontrolní skupiny s 95% intervaly spolehlivosti (CI), Z-skóry a Crawfordovým t-testem porovnávajícím každého pacienta s její příslušnou kontrolní skupinou.
Pacient ML
Účastníci kontroly: Průměrné hodnoty mediánů RT se u odpovědí na levý a pravý cíl nelišily (t = 0,86. Obrázek 7 ukazuje, že RT byly kratší (RTE) pro bilaterální cíle než pro levé (t = 4,7, p = 0,001, nebo pravé (t1,11) = 7,8, p < 0,001) unilaterální cíle.

OBRÁZEK 7. Obrázek 7 ukazuje, že RT byly kratší (RTE) pro bilaterální cíle než pro pravé cíle. Mediány RT u pacientů ML a průměr mediánů RT u účastníků kontrolní skupiny pro cíle objevující se v levém zorném poli, v pravém zorném poli a současně v obou zorných polích. Chybové sloupce pro účastníky kontrolní skupiny označují standardní chyby.
Medián RT pro pokusy s bilaterálními cíli byl odečten od průměru mediánu RT pro levé a pravé unilaterální cíle, aby bylo možné vypočítat RTE pro každého účastníka kontrolní skupiny.
Skupinový průměr RTE byl 33 ms (SEM = 6,65).
Pacient ML měl medián RT delší než kontrolní skupina (obr. 7) a byl nad horní hranicí 95% CI průměrného RT kontrolní skupiny pro všechny tři podmínky (levá = 322 ms; pravá = 307 ms; obě = 277 ms). RT na jednostranné cíle v pravém (kontralezionálním) poli byly delší než u jednostranných cílů v levém (ipsilezionálním) poli (t = 4,9, p = 0,001). Nebyl zaznamenán žádný RTE. RT pro bilaterální cíle (361 ms) nebyly kratší než pro unilaterální cíle v ipsilesionálním (levém) zorném poli (363 ms). Tabulka 1 (dole) ukazuje, že RTE vypočtené pro pacientku ML byly významně kratší než RTE pro její kontrolní skupinu.
Diskuse
U dvou pacientů, u nichž byl brachium SC poškozen v jedné hemisféře, byly měřeny jednoduché RT pro detekci jednotlivých cílů v kontralezionálním nebo ipsilezionálním zorném poli a bilaterálních cílů. U jednoho z pacientů (mladá žena) se poškození nacházelo v levé hemisféře a bylo způsobeno traumatickým poraněním mozku; a přestože léze zahrnovala dolní koliculus a zasahovala do tegmenta středního mozku, poškození SC jako takového nebylo pomocí neurozobrazení s vysokým rozlišením patrné. U druhého pacienta (starší žena) se poškození nacházelo v pravé hemisféře a bylo způsobeno hypertenzním krvácením, které rovněž způsobilo rozsáhlé poškození mediálního pulvina a laterálního talamu a zasahovalo do dorzálního středního mozku včetně rostrálního SC. U obou pacientů byl jednoduchý RT k detekci zrakových cílů v kontralezionálním poli zpomalen ve srovnání s ipsilezionálním polem. Ani u jednoho z pacientů se neprojevil efekt prostorové sumace (RTE) při detekci cíle, když byly podněty prezentovány do obou zrakových polí.
Tato pozorování vrhají světlo na roli SC při detekci cíle a konkrétněji na roli zrakových aferentů do SC. Posner et al. (1980) operačně definovali detekci jako projev schopnosti provést libovolnou reakci na vizuální signál (např. jednoduché stisknutí klávesy prstem, jako v tomto experimentu). Předpokládali, že detekce vyžaduje alokaci pozornosti pro výběr cíle ke zpracování v systému s omezenou kapacitou, který mu dává prioritu, aby na něj bylo možné působit.
Song et al (2011) ukázali, že inaktivace opičí SC způsobuje nápadné deficity výběru cíle, které nelze snadno vysvětlit jako prosté narušení zrakového vnímání nebo motorického provedení, a navrhli, že přispívá k obecnějšímu účelu mapy priorit. V tomto experimentu byly opicím předloženy dva podněty v protilehlých zorných polích a opice měly pomocí odpovědi na dosah ruky určit, který z nich se objeví jako první. Jedna opice byla trénována, aby dosáhla na podnět, který se objevil jako první, a jedna opice byla trénována, aby dosáhla na cíl, který se objevil jako druhý. V obou případech byla zjištěna silná tendence k výběru cíle, který se nacházel v zorném poli kontralaterálně od léze. Vzhledem k tomu, že stejné zkreslení bylo pozorováno i u opice, která dosáhla na cíl, který se objevil jako druhý, nemohlo být zkreslení způsobeno zpožděním při vnímání kontralezionálního cíle. Kolikulární inaktivace nezpůsobila zhoršení percepčního úsudku o tom, který podnět se objevil jako první. Spíše byly podněty v zorném poli kontralaterálně od inaktivovaného koliculu znevýhodněny v tom, že byly upřednostněny pro akci.
Zhaoping (2016) nedávno zdůraznil rozdíl mezi neuronálními mapami saliencí a mapami priorit a argumentoval pro evoluční migraci percepční mapy saliencí z optického tekta/SC do primární zrakové kůry. Navrhla, že signály salience mohou být přenášeny do prioritní mapy v SC, aniž by se do ní promítala vlastnost ladění rysů.
Brachium SC přenáší aferentní zrakové signály do povrchových vrstev koliculu ze sítnice prostřednictvím retinotektálního traktu; většina zrakových aferentních vláken přenášených brachiem jsou však projekce z primární zrakové kůry. Pokud je brachium narušeno, mohou být salience signály z primární zrakové kůry přenášeny do colliculu přes frontální oční pole. Pokud však chybí přímé projekce do povrchových vrstev koliculu ze sítnice nebo ze zrakové kůry, jak předpokládáme u pacientů s lézí brachia, trvá aktivace koliculární mapy priorit déle, což má za následek pomalejší reakce na kontralaterální zrakové signály.
Jak bylo uvedeno v úvodu, příspěvek k RTE, který je založen na pravděpodobnostním modelu dostihového koně, závisí na detekci obou cílů; a na dostihu, v němž je účinnost nezávislých kanálů, v nichž jsou oba cíle přenášeny, rovnocenná, takže výsledek dostihu na dosažení prahu detekce je od pokusu k pokusu náhodný. Vzhledem k tomu, že práh detekce kontralezionálních signálů je u zde uváděných pacientů vyšší, nemohly by redundantní cíle přinášet prospěch na základě stochastického dostihu; to znamená, že výsledek závodu mezi ipsilesionálními a kontralezionálními signály není náhodný – kontralezionální signály mají větší pravděpodobnost, že závod prohrají, než že ho vyhrají.
Výzkum u hemianopických pacientů nicméně ukázal, že k RTE přispívá také nervová sumační složka, která nezávisí na detekci obou cílů (Marzi et al., 1986; Tomaiuolo, 1997; de Gelder et al., 2001). Vzhledem k tomu, že u pacientů s komisurotomií byla prokázána RTE(Marzi et al., 1986; Reuter-Lorenz et al., 1995), bylo argumentováno, že interhemisférická integrace signálů přes vertikální meridián musí probíhat subkortikálně.
Savazzi a Marzi (2004) ukázali, že k nervové sumaci nedochází při krátkovlnných chromatických cílových podnětech ani u neurologicky intaktních lidí, ani u pacientů s rozděleným mozkem. Leh et al (2006b) uvedli, že zatímco u některých pacientů s hemianopickou hemisférektomií docházelo k RTE u achromatických podnětů, u těchto pacientů nedocházelo k RTE, pokud byly jako cíle použity chromatické podněty o krátké vlnové délce. Krátkovlnné (tj. fialové) podněty aktivují pouze S-konečky v sítnici. Gangliové buňky sítnice, které dostávají vstup z S-konečků, se nepromítají ani přímo do SC, ani do magnocelulárních vrstev laterálního genikulárního jádra (De Monasterio, 1978). Vzhledem k tomu, že projekce z primární zrakové kůry do SC přes brachium SC přenášejí zrakové signály pouze z magnocelulárních genikulostriátních aferentů (Schiller et al., 1979), selhání S-konečných podnětů při vyvolání RTE naznačuje, že nervová sumace probíhá v SC a je závislá na zrakových aferentech přenášených přes jeho brachium.
U pacientů s rozděleným mozkem, hemianopických pacientů se slepotou a z experimentů využívajících podněty s krátkou vlnovou délkou tedy existují sbíhající se důkazy, že k nervové sumaci dochází v SC. Nicméně i když podněty o krátké vlnové délce neaktivují gangliové buňky sítnice, které se promítají do SC přes brachium, nelze z toho vyvodit závěr, že colliculus je vůči těmto podnětům zcela slepý. Záznamy jednotlivých jednotek v opičím SC prokázaly, že krátkovlnné podněty aktivují odpovědi v povrchových vrstvách koliculu. Kromě toho traktografie prokázala u pacientů po hemisférektomii s RTE (tj. slepotou), ale ne u pacientů, kteří nevykazovali známky slepoty, že SC má spojení s intaktní hemisférou (Leh et al..), 2006a).
Zjištění současného šetření tedy přinášejí první přímý důkaz, že v SC dochází k prostorové sumaci, která je závislá na přenosu zrakových signálů přes jeho raménko.
Etické prohlášení
Tato studie byla provedena v souladu se směrnicemi a se schválením protokolu etickou komisí NHS, Bangor, UK, a etickou komisí School of Psychology University, Bangor, UK. Od všech účastníků byl získán ústní a písemný informovaný souhlas v souladu s Helsinskou deklarací.
Příspěvky autorů
MvK: Koncipoval výzkum, navrhl a naprogramoval experiment, shromáždil a analyzoval údaje pacientů a podílel se na napsání rukopisu. KK: Shromáždil kontrolní údaje, prostudoval příslušnou literaturu, analyzoval kontrolní údaje a přispěl k napsání rukopisu. RR: Provedl nábor pacientů a analyzoval neurozobrazovací studie včetně virtuální disekce s traktografií DTI, připravil anamnézy pacientů a podílel se na psaní rukopisu.
Prohlášení o střetu zájmů
Autoři prohlašují, že výzkum byl prováděn bez jakýchkoli komerčních nebo finančních vztahů, které by mohly být chápány jako potenciální střet zájmů.
Behrens, T., Woolrich, M. W., Jenkinson, M., Johansen-Berg, H., Nunes, R. G., Clare, S. a další (2003). Charakterizace a šíření nejistoty v difuzně váženém MR zobrazení. Magn. Reson. Med. 50, 1077-1088. doi: 10.1002/mrm.10609
PubMed Abstract | CrossRef Full Text | Google Scholar
Behrens, T. E., Berg, H. J., Jbabdi, S., Rushworth, M. F. a Woolrich, M. W. (2007). Pravděpodobnostní difuzní traktografie s více orientacemi vláken: co můžeme získat? Neuroimage 34, 144–155.
Google Scholar
de Gelder, B., Pourtois, G., van Raamsdonk, M., Vroomen, J., and Weiskrantz, L. (2001). Unseen stimuli modulate conscious visual experience: evidence from interhemispheric summation. Neuroreport 12, 385–391. doi: 10.1097/00001756-200102120-00040
CrossRef Full Text | Google Scholar
De Monasterio, F. M. (1978). Properties of ganglion cells with atypical receptive-field organization in retina of macaques. J. Neurophysiol. 41, 1435–1449.
PubMed Abstract | Google Scholar
Fries, W. (1984). Cortical projections to the superior colliculus in the macaque monkey: a retrograde study using horseradish peroxidase. J. Comp. Neurol. 230, 55–76. doi: 10.1002/cne.902300106
PubMed Abstract | CrossRef Full Text | Google Scholar
Georgy, L., Celeghin, A., Marzi, C. A., Tamietto, M., and Ptito, A. (2016). Horní kolikulus je citlivý na konfiguraci podnětů podobnou gestům u pacientů po hemisférektomii. Cortex 81, 151-161. doi: 10.1016/j.cortex.2016.04.018
PubMed Abstract | CrossRef Full Text | Google Scholar
Hershenson, M. (1962). Reakční čas jako míra intersenzorické facilitace. J. Exp. Psychol. 63, 289-293. doi: 10.1037/h0039516
CrossRef Full Text | Google Scholar
Hikosaka, O. a Wurtz, R. (1983). Zrakové a okulomotorické funkce opičí substantia nigra pars reticulata. III. paměťově podmíněné zrakové a sakádové odpovědi. J. Neurophysiol. 49, 1268-1284.
PubMed Abstract | Google Scholar
Leh, S. E., Johansen-Berg, H. a Ptito, A. (2006a). Nevědomé vidění: nové poznatky o neuronálním korelátu slepého vidění pomocí difuzní traktografie. Brain 129(Pt 7), 1822–1832.
PubMed Abstract | Google Scholar
Leh, S. E., Mullen, K. T., and Ptito, A. (2006b). Absence of S-cone input in human blindsight following hemispherectomy. Eur. J. Neurosci. 24, 2954–2960.
PubMed Abstract | Google Scholar
Marzi, C., Tassinari, G., Aglioti, S., and Lutzemberger, L. (1986). Spatial summation across the vertical meridian in hemianopics: a test of blindsight. Neuropsychologia 24, 749–758. doi: 10.1016/0028-3932(86)90074-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Miller, J. (1982). Divided attention: evidence for coactivation with redundant signals. Cogn. Psychol. 14, 247–279. doi: 10.1016/0010-0285(82)90010-X
CrossRef Full Text | Google Scholar
Miniussi, C., Girelli, M., and Marzi, C. A. (1998). Neural site of the redundant target effect electrophysiological evidence. J. Cogn. Neurosci. 10, 216–230. doi: 10.1162/089892998562663
CrossRef Full Text | Google Scholar
Pare, M., and Wurtz, R. (1997). Monkey posterior parietal cortex neurons antidromically activated from superior colliculus. J. Neurophysiol. 78, 3493–3497.
PubMed Abstract | Google Scholar
Poliva, O., Bestelmeyer, P. E., Hall, M., Bultitude, J. H., Koller, K., and Rafal, R. D. (2015). Funkční mapování lidské sluchové kůry: fMRI vyšetření pacienta se sluchovou agnózií v důsledku úrazu dolního koliculu. Cogn. Behav. Neurol. 28, 160-180. doi: 10.1097/WNN.0000000000000072
PubMed Abstract | CrossRef Full Text | Google Scholar
Posner, M., Snyder, C., and Davidson, B. (1980). Pozornost a detekce signálů. J. Exp. Psychol. Gen. 109, 160-174. doi: 10.1037/0096-3445.109.2.160
CrossRef Full Text | Google Scholar
Raab, D. H. (1962). Statistické usnadnění jednoduchých reakčních časů. Trans. N. Y. Acad. Sci. 24, 574-590. doi: 10.1111/j.2164-0947.1962.tb01433.x
CrossRef Full Text | Google Scholar
Reuter-Lorenz, P., Nozawa, G., Gazzaniga, M. S., and Hughes, H. C. (1995). Fate of neglected targets: a chronometric analysis of redundant target effects in the bisected brain (Osud zanedbaných cílů: chronometrická analýza nadbytečných cílových efektů v rozpolceném mozku). J. Exp. Psychol. Hum. Percept. Perform. 21, 211-230. doi: 10.1037/0096-1523.21.2.211
PubMed Abstract | CrossRef Full Text | Google Scholar
Savazzi, S., and Marzi, C. A. (2004). The superior colliculus subserves interhemispheric neural summation in both normalals and patients with a total section or agenesis of the corpus callosum. Neuropsychologia 42, 1608-1618. doi: 10.1016/j.neuropsychologia.2004.04.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Schiller, P. H., Malpeli, J. G., and Schein, S. J. (1979). Composition of geniculostriate input ot superior colliculus of the rhesus monkey. J. Neurophysiol. 42, 1124-1133.
PubMed Abstract | Google Scholar
Sommer, M. a Wurtz, R. (2000). Složení a topografická organizace signálů vysílaných z čelního očního pole do horního koliculu. J. Neurophysiol. 83, 1979–2001.
PubMed Abstract | Google Scholar
Song, J. H., Rafal, R. D., and McPeek, R. M. (2011). PNAS Plus: deficits in reach target selection during inactivation of the midbrain superior colliculus. Proc. Natl. Acad. Sci. U.S.A. 108, E1433–E1440. doi: 10.1073/pnas.1109656108
PubMed Abstract | CrossRef Full Text
Tomaiuolo, F. (1997). Blindsight in hemispherectomized patients as revealed by spatial summation across the vertical meridian. Brain 120, 795–803. doi: 10.1093/brain/120.5.795
PubMed Abstract | CrossRef Full Text | Google Scholar
Zhaoping, L. (2016). From the optic tectum to the primary visual cortex: migration through evolution of the saliency map for exogenous attentional guidance. Curr. Opin. Neurobiol. 40, 94–102. doi: 10.1016/j.conb.2016.06.017
PubMed Abstract | CrossRef Full Text | Google Scholar