Existují rodokmeny a populace, kde je princip panmiktického páření v podstatě porušen. Obvykle se jedná o geograficky nebo sociálně relativně izolovaná malá společenství. V těchto společenstvích může být široce rozšířen inbreeding, tj. příbuzenské páření různých řádů (rodič – dítě, sourozenci, bratranci, strýc – neteř atd.).
Problém vlivu inbreedingu na populaci je důležitým problémem genetiky 1. Nyní tento problém ještě nezískal konečné řešení.
Ve většině případů se příbuzenská plemenitba v rodokmenu zohledňuje na třech generacích. Jedná se o nejvzdálenější příbuzné, kteří přicházejí v úvahu při příbuzenském křížení existuje bratranec a sestřenice z druhého kolena.
K příbuzenskému křížení se přihlíží pomocí faktoru příbuzenského křížení.
Faktor příbuzenského křížení je pravděpodobnost, že dvě alely jsou shodné podle původu.
Uvažujme metodu výpočtu faktoru příbuzenského křížení v rodokmenu, kterou nabízí Wright 1, 2.
Pro zjištění faktoru příbuzenského křížení je nutné:
zjistit pro snoubící se potomky všechny cesty v rodokmenu, které přes společného předka spojují tyto potomky.
počítat počet kroků (střídání generací) na každé cestě.
Poté je faktor inbreedingu roven:
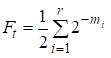 ……(1)
……(1)
kde r je počet cest, m1, m2, …. mi- počet kroků na každé cestě.
Znovu si všimneme, že počet sčítanců v součtu (1) je roven počtu cest, které přes společné předky spojují sňatky.
Uvažujme konkrétní případy rodokmenů, obr. 1. Na tomto obrázku čtverec znamená mužského jedince kruh – ženského jedince. Tečkované čáry a shodné číslice udávají možné způsoby v rodokmenu, které přes společného předka (muže nebo ženu) spojují potomky příbuzenské sňatky. Na některých místech způsobů, obrázek 1с, jsou dvě číslice 1 a 2. To určuje, že taková místa jsou společná pro různé způsoby.
Sňatek otec – dcera je incest, obrázek 1а. Potomek (žena) je zde spojen s předkem (mužem) 1 cestou s jedním krokem, tj. m = 1. Existuje tedy faktor příbuzenské plemenitby
Ft = 1/2,2-1 = 1/4
Existuje sňatek bratr – sestra (sibs), obrázek 1b. Číslice udávají počet způsobů. Počet stejných číslic na cestě od ženatého muže k ženě je roven počtu kroků.
Nechť každý potomek, který se oženil, spojí s partnerkou dvě cesty se dvěma kroky na každé cestě, tj. m = 2. Existuje tedy faktor inbreedingu.
Ft = 1/2.(2-2 + 2-2) = 1/4
Sňatek mezi bratranci a sestřenicemi je znázorněn na obrázku 1с.
Obrázek 1s. Některé druhy příbuzenského křížení v rodokmenu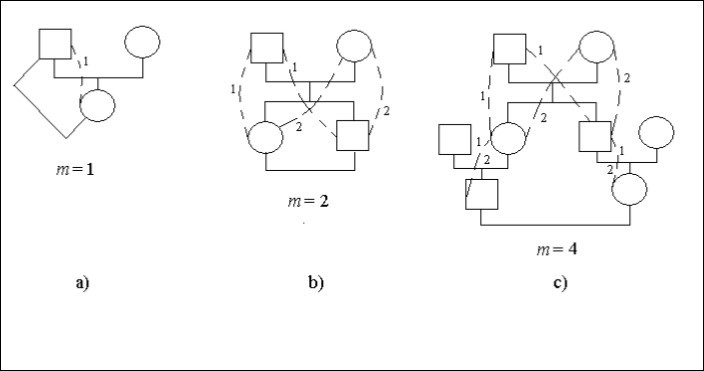
Každý potomek, který uzavřel sňatek, se spojí s partnerem 2 způsoby po 4 krocích na každém způsobu, tj. m = 4. V případě, že se bratranec nebo sestřenice oženil nebo vdala, je příbuzenský vztah mezi nimi ukončen. Existuje tedy faktor inbreedingu.
Ft = 1/2.(2-4 + 2-4) = 1/16
Vzorec (1) je snadno pochopitelný z následující logiky.
Zřejmě je 1/2 genů dítěte společná s každým rodičem, 1/4 – s dědečkem a babičkou, 1/8 – s pradědečkem a prababičkou atd. Pokud od ženy k nějakému předkovi vede cesta s η kroky, podíl společných genů s tímto předkem je roven 2-η. Pokud od muže k jakémukoli předkovi vede cesta s μ kroky, je podíl společných genů roven 2-μ. Při sňatku muže a ženy se společným předkem bude podíl společných genů s tímto předkem roven 2-η. 2-μ = 2-m, kde m = η + μ je celkový počet kroků přes společného předka od ženy až po sňatek s mužem. Velikost 1/2 2-m udává pravděpodobnost, že gen náhodně vybraný u ženy bude svým původem shodný s genem náhodně vybraným u muže. Stejná velikost udává příspěvek k faktoru inbreedingu Ft jednou cestou přes společného předka od ženy až po sňatek s mužem.
Příklad pokud bude společný předek vzdálen od sňatku na η = μ = 5 generací, pravděpodobnost, že budou mít společný gen, je stejná. 1/2 2-(5+5) = 1/2048 Tuto velikost při analýze páření lze zanedbat. (Tabulka 1)
V tabulce 1.jsou navíc uvedeny faktory inbreedingu pro některá příbuzenská manželství.
| Druh příbuzenského sňatku. manželství | Faktor příbuzenské plemenitby Ft |
| Druhý bratranec a sestřenice | 1/32 |
| Bratranec strýc -. niece | 1/32 |
| Third cousins | 1/64 |
The inbreeding factor Ft is the probability of two alleles in posterity are identical by origin i.e. occur from same gene of ancestor 1. Or, that too most, the inbreeding factor is a probability of the allele origin in posterity is consanguineous. Hence, the probability of the non-consanguineous origins of identical alleles in homologous chromosomes is equal 1 – Ft.
Let’s find total probability of the recessive homozygote aa occurrence (eventаа) at random mating for an autosomal genes 3:
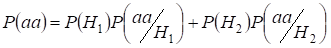 …….(2)
…….(2)
This formula follows from the following logic of mating: Původ shodných alel v homologních autosomech je nekonsangvinní (pravděpodobnost této hypotézy je P(H1) = 1- Ft )vzniká také homozygotаа (podmíněná pravděpodobnost této události je P(aa/H1) = q2) nebo původ shodných alel je konsangvinní (pravděpodobnost této hypotézy je P(H2) = Ft ) a vzniká homozygotаа (podmíněná pravděpodobnost této události je P(aa/H2) = q). Pravděpodobnost q výskytu homozygota aa v druhém případě je dána tím, že pokud je alela a v jednom autosomu, pak při konsanguinním původu homozygota by stejná alela nutně měla být i v homologním autosomu.
Z toho vyplývá, že celková pravděpodobnost výskytu recesivního homozygota je rovna:
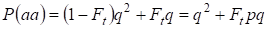 ……….(3)
……….(3)
kde se bere v úvahu p = 1 – q, tam je frekvence dominantní alely a .
Podobně tam najdeme celkovou pravděpodobnost výskytu dominantního homozygota AA při náhodném páření:
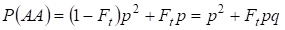 ……….. (4)
……….. (4)
Podle (3) a (4) se tedy celková frekvence homozygotů (dominantních i recesivních) v inbredním rodokmenu zvyšuje na 2Ftpq. Proto se frekvence heterozygotů při stejné velikosti snižuje a stává se rovnou 2pq – 2pqFt = 2pq(1-Ft), protože součet frekvencí všech zygot je roven 1.
Vzhledem k faktoru inbreedingu a také (3) a (4) Hardyho – Weinbergova zákona v následující generaci inbredního rodokmenu pro autozomální genom je třeba zapsat rozdělení genotypů zygot jako 4:
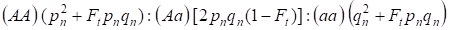 …………(5)
…………(5)
Kde pn je frekvence dominantních alel A v generaci n, qn – frekvence recesivních alel a, takže pn + qn = 1.
Poměr (5) odkazuje také na Wrightův poměr 5. Wrightův poměr ukazuje, že v inbredním rodokmenu po konsanguinním páření jedinců jsou pozorovány následující efekty: dochází ke zvýšení podílu autozomálních homozygotů v rodokmenu na Ft pnqna snížení podílu autozomálních heterozygotů v (1-F1) ve srovnání s panmiktickou populací.
Ale již přes jednu generaci se obnovuje Hardyho – Weinbergova rovnováha, která určuje malý význam samostatného konsanguinního páření pro rodokmen:
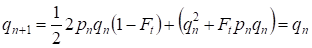 ………(6)
………(6)
Zřejmě pro populaci jako celek bude mít samostatný akt konsanguinního páření ještě menší význam než pro rodokmen.
Stejný vliv variability frekvence genotypů zygoty je pozorován i pro geny vázané na pohlaví.
Rozložení genotypů vázaných na chromozom H v panmiktickém rodokmenu popisuje Hardyho – Weinbergův zákon jako 1:
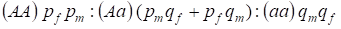 ………..(7)
………..(7)
Frekvenci dominantní alely A budeme označovat u mužů pm a u žen pf. Pro recesivní alelya je to obdobně qm a qf .
Při páření existuje poměr genotypů u žen podle součinu (pf + qf) (pm +qm).
Najděme celkovou pravděpodobnost výskytu recesivního homozygota aa při náhodném páření pro geny dcery spojené s chromozomemХ podle vzorce (2). Logika událostí je podobná jako v případě autozomů s tím rozdílem, že podmíněná pravděpodobnost výskytu genotypuаа z nesourodých alel podle (7) je rovna P(aa/H1) = qmnqfn. Pro konsanguinní alely je podmíněná pravděpodobnost výskytu genotypuаа rovna P(aa/H2) = qf(n+1). U dcer, tj. v generaci n+1, je pravděpodobnost výskytu homozygota aa dána tím, že pokud je alela a v jednom chromozomu H, pak při konsanguinním původu homozygota by stejná alela nutně měla být i v homologním chromozomu H.
Z toho vyplývá, že celková pravděpodobnost výskytu recesivního homozygota je rovna:
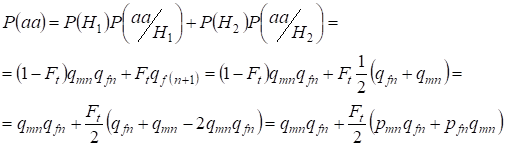 ……(8)
……(8)
kde pmn je frekvence dominantní alely A v generaci n u mužů a pfn – u žen. Pro recesivní alely a odpovídající qmn a qfn . Kromě toho se používá qmn =1 – qmn a pfn = 1 – qfn.
Podobně zjistíme celkovou pravděpodobnost výskytu dominantního homozygota pomocí:
 ,
,
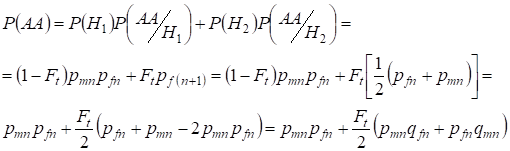 ……….(9)
……….(9)
a heterozygota P(Aa) = 1- P(aa) – P(AA) zjistíme rozložení genotypů v chromozomech H inbredního rodokmenu při páření:
 ……..(10)
……..(10)
Současně se ukáže, že stejně jako u autozomů jsou inbrední přídavky v dominantních a recesivních frekvencích homozygotů v pohlavně vázaných chromozomech shodné.
Jiný způsob zjištění poměru (10) bude spočívat v následujícím:
Najdeme nejprve celkovou pravděpodobnost výskytu heterozygota Aa, tedy P(Aa) při příležitosti náhodného příbuzenského páření. Tato pravděpodobnost vyplývá z následující logiky páření: při nesourodém původu alel (pravděpodobnost této hypotézy je P(H1) = 1 – Ft ) vzniká také heterozygot A Aa (podmíněná pravděpodobnost této události je P(Aa/H1) = pmnqfn + pfnqmn, viz vzorec (7)) nebo je původ alel příbuzenský (pravděpodobnost této hypotézy je P(H2) = Ft) a vzniká heterozygot Aа (podmíněná pravděpodobnost této události je rovna nule P(Aa/H2) = 0 ). Poslední tvrzení je definováno tím, že při konsanguinním původu nemohou podobné alely v homologních chromozomech Х vzniknout heterozygotАа složený z různých alel.
Takže pomocí vzorce celkové pravděpodobnosti zjistíme celkovou pravděpodobnost výskytu heterozygota Aa:
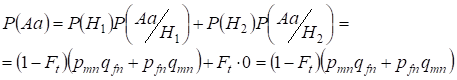 ……..(11)
……..(11)
Snížení frekvence heterozygotů Aa nastává, jak bylo prokázáno dříve, v důsledku shodného zvýšení frekvence dominantních homozygotů AA a recesivních homozygotů aa. Například zvýšení frekvence recesivních homozygotů aa je stejné:
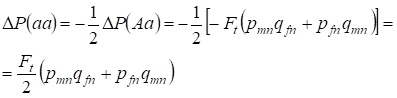 ……..(12)
……..(12)
kde snížení frekvence heterozygotů Aa vzhledem k panmiktickému rodokmenu podle (7) a (11) je stejné. ΔP(Aa) = – Ft (pmnqfn + pfnqmn) Pomocí (11) a (12) k aktualizaci (7) pro případy inbredního křížení zjistíme rozložení genotypů (10) v chromozomech H v dceřiném inbredním rodokmenu.
V následující generaci se frekvence recesivních alel a u žen rovná:
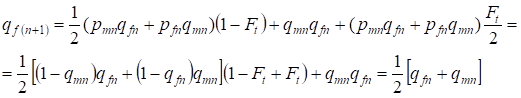 ……..(13)
……..(13)
To plně odpovídá stejné frekvenci při absenci inbreedingu, tj. dochází k obnovení Hardyho – Weinbergovy rovnováhy nebo je přesnější přiblížení k rovnováze.
Z poměru (7) a (13) vyplývá, že jako celek se frekvence alel v rodokmenu na konto inbreedingu nemění. Dochází pouze k přerozdělení alel. Část alel opouští heterozygoty a přechází na homozygoty.