1. Cilium
2. Flagelo
3. Estructura
4. Movimiento
5. Formación
Los microtúbulos son componentes del citoesqueleto con importantes funciones en la fisiología celular. El andamiaje microtubular del citoplasma es altamente plástico gracias a la capacidad de polimerización y despolimerización de los microtúbulos. Sin embargo, no todos los microtúbulos de las células se encuentran en esta fase de contracción o crecimiento. Los cilios, los flagelos y los centríolos/cuerpos basales son estructuras celulares que contienen microtúbulos muy estables (en número y longitud) y muy organizados. En esta página nos ocupamos de los cilios y los flagelos.
1. Cilio
Los cilios son protuberancias celulares delgadas y largas de unos 0,25 µm de diámetro y de unos 10 a 15 µm de longitud, que pueden encontrarse en las células animales y en algunas especies eucariotas unicelulares. Suelen estar fuertemente empaquetados en la superficie libre de las células epiteliales (Figuras 1 y 2), como el epitelio de las vías respiratorias, el epitelio de los conductos reproductores, las branquias de peces y bivalvos, etcétera. Los cilios son estructuras móviles y su función principal es mover el líquido circundante, como el moco de la superficie del tracto respiratorio, el agua alrededor del epitelio de las branquias, pero también el ovocito en el conducto de Falopio femenino. Muchos organismos unicelulares pueden moverse impulsados por los cilios, y otros pueden utilizarlos para generar remolinos de agua para atrapar el alimento. Los cilios nodales del embrión han sido implicados en la iniciación del eje izquierda-derecha durante el desarrollo embrionario de los vertebrados. El movimiento de los cilios es como un batido que impulsa el líquido paralelo a la superficie celular.
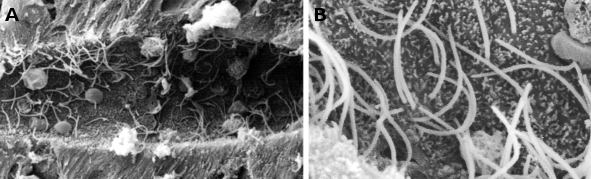
Figura 1. Imágenes de microscopía electrónica de barrido que muestran el canal central de una médula espinal de lamprea. Se observan muchos cilios (a mayor aumento en B) y pequeñas microvellosidades en el dominio apical de las células ependimarias.
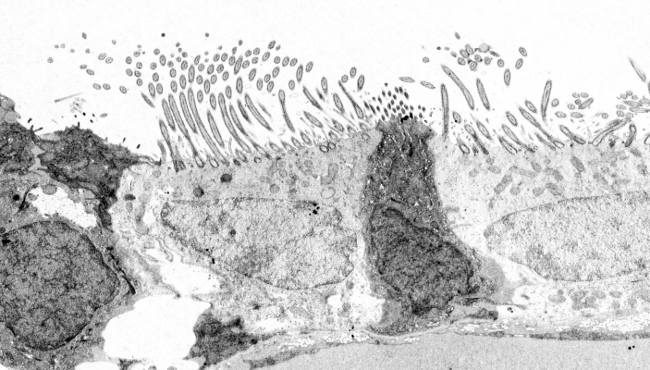
Figura 2. Imágenes de microscopía electrónica de transmisión del epitelio respiratorio. Las células con citoplasma claro muestran muchos cilios en su superficie apical.
Hay cilios que no pueden moverse, y por tanto no están destinados al movimiento de líquidos. Estos cilios se conocen como cilios primarios. La mayoría de las células estudiadas hasta ahora (exceptuando los glóbulos rojos) llevan cilios primarios: células del oviducto, neuronas, condrocitos, células del ectodermo, células mesenquimales, células epiteliales urinarias, hepatocitos e incluso células cultivadas. Inicialmente, se pensaba que los cilios primarios eran cilios no funcionales. Sin embargo, se encontraron muchos tipos de receptores y canales iónicos en la membrana ciliar, por lo que se consideraron estructuras sensoriales celulares. Por ejemplo, los receptores olfativos se encuentran en los cilios de sus dendritas, y los segmentos externos de los bastones y conos de la retina son en realidad cilios modificados. Algunos receptores están más empaquetados en la membrana ciliar que en otros dominios de la membrana plasmática. Además, hay una gran variedad de moléculas en el interior de los cilios que participan en funciones de transducción de señales. La mayor relación superficie/volumen de un cilio hace que las respuestas moleculares intraciliares sean más intensas y eficientes que si estuvieran fuera del cilio. Además de la señal química, los cilios primarios pueden detectar el movimiento de fluidos fuera de la célula y funcionar como mecanorreceptores.
2. Flagelos
Los flagelos son similares a los cilios, pero son mucho más largos, de unos 150 µm de longitud, y ligeramente más gruesos. Son bastante menos numerosos que los cilios en las células. La función principal de los flagelos es mover la célula. El movimiento del flagelo es diferente al del cilio porque la dirección del movimiento es perpendicular a la superficie de la célula (no paralela), es decir, la dirección del eje longitudinal del flagelo. Los flagelos se observan con frecuencia en células móviles como los organismos unicelulares y los espermatozoides.
Estructura
Los cilios y los flagelos son estructuras complejas que contienen más de 250 proteínas diferentes. Ambos comparten la misma organización central de microtúbulos y otras proteínas asociadas, conocidas en conjunto como axonema, y limitadas por la membrana plasmática (Figura 3). Además del axonema, hay muchas moléculas solubles dentro de los cilios/flagelos que constituyen la matriz. El axonema está formado por 9 pares de microtúbulos alrededor de otro par central de microtúbulos. Esta organización se puede escribir como (9 x 2) + 2. Los cilios primarios carecen de un par central de microtúbulos. Cada microtúbulo del par central está formado por 13 protofilamentos, pero los microtúbulos de los pares periféricos comparten algunos protofilamentos entre sí. Así, un par periférico está formado por los microtúbulos A y B. El microtúbulo A contiene 13 protofilamentos y el microtúbulo B contiene 10 u 11 protofilamentos, compartiendo 2 o 3 con el microtúbulo A.

Figura 3. Principales componentes moleculares de los cilios y los flagelos. En los cilios primarios, el par central está ausente.
La organización de los microtúbulos del axonema es el resultado de un andamiaje de proteínas. Ya se han encontrado doce proteínas como constituyentes del axonema que participan en el mantenimiento de la organización de los microtúbulos. Los pares de microtúbulos periféricos vecinos están conectados entre sí por la nexina. En cada par, el microtúbulo A está conectado por radios proteicos a un anillo central que contiene el par o los microtúbulos centrales. La dineína es una proteína motora asociada a los microtúbulos periféricos que participan en el movimiento de los cilios y los flagelos.

Figura 4. Ultraestructura de un cilio de una célula ependimaria de la médula espinal. (9+2)x2 significa 9 pares periféricos y 1 par central de microtúbulos.
Los microtúbulos se polimerizan a partir de los cuerpos basales (Figuras 3 y 4). Los cuerpos basales están formados por 9 microtúbulos triples que forman un cilindro (similar a los centríolos). Carecen de un par central de microtúbulos, por lo que es (9×3)+0. En cada triplete, sólo un microtúbulo (microtúbulo A) tiene un conjunto completo de protofilamentos, mientras que los microtúbulos B y C comparten algunos de ellos entre sí. Desde el cuerpo basal, los microtúbulos A y B crecen y forman los microtúbulos periféricos del axonema. Justo por encima del cuerpo basal, hay una región de los cilios conocida como zona de transición que contiene los 9 pares periféricos y ningún par central. Inmediatamente después de la zona de transición, se encuentra la placa basal, a partir de la cual se polimeriza el par central de microtúbulos para completar el axonema. Todos los microtúbulos tienen el extremo positivo hacia la punta del cilio/flagelo. El extremo proximal del cuerpo basal (el interno, o extremo negativo de los microtúbulos) está anclado al citoesqueleto celular a través de largas fibras proteicas llamadas raicillas ciliares
Además del axonema, los cilios/flagelos tienen otros compartimentos. La membrana contiene muchos receptores y canales para percibir el entorno, especialmente en los cilios primarios. La fase fluida del interior se denomina matriz, que ayuda a mantener organizada toda la estructura y se encarga de transducir la información recogida por los receptores de la membrana. Otras zonas diferenciadas son el cuerpo basal situado en la base y la parte apical del cilio/flagelo, que contiene proteínas que estabilizan los extremos positivos de los microbulos.
4. Movimiento
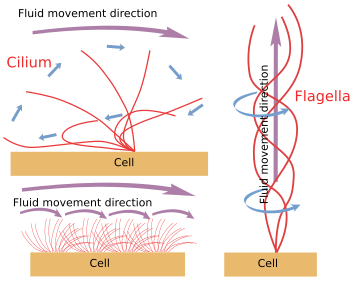
Figura 6. Modelos de movimiento de cilios y flagelos. Generan diferentes direcciones de movimiento del fluido.
Cuando los cilios/flagelos se desprenden mecánicamente de la célula, siguen moviéndose hasta que se agota el almacén de ATP. Esto significa que el mecanismo de movimiento es intrínseco (Figura 6). En realidad, el movimiento se produce por el deslizamiento de un par periférico de microtúbulos sobre el vecino. Las proteínas nexina y spoke impiden la desorganización del axonema, pero permiten estos movimientos. La dineína es la proteína motora responsable del movimiento de deslizamiento. La energía la proporciona el ATP. Las dineínas se anclan con su parte globular al microtúbulo A de un par periférico y con su parte de cola al microtúbulo B del par contiguo. El mecanismo molecular es similar al de las dineínas citosólicas, pero en lugar de transportar una carga, mueve un microtúbulo. Para que el movimiento sea eficaz, se necesita una coordinación de las dineínas del axonema. Las ondas de calcio en el interior de los cilios/flagelos pueden coordinar la activación de las dineínas y cambiar la frecuencia de movimiento cuando sea necesario. Es de notar que no todas las dineínas tienen que activarse al mismo tiempo, sino en sincronía.
5. Formación
Durante la diferenciación, las células producen todos los cilios y flagelos necesarios para su fisiología normal. Esto significa que todos ellos deben ser generados desde cero. Los microtúbulos del axonema se nuclean a partir de los microtúbulos A y B de los cuerpos basales, por lo que se necesita un cuerpo basal por cilio/flagelo. ¿Cómo se forman los cuerpos basales múltiples? Hay al menos tres formas de producir cuerpos basales: a) utilizando los centriolos como plantillas para nuclearlos; b) a partir de un material amorfo conocido como deuterosoma; c) en las plantas, hay agregados proteicos distintos que pueden nuclear cuerpos basales.
Numerosas patologías humanas son consecuencia de fallos en los cilios/flagelos. Se conocen como ciliopatías, e incluyen la lateralidad aleatoria, el cierre incorrecto del tubo neural, la polidactilia, las patologías quísticas del riñón, el hígado y el páncreas, la degeneración de la retina, la obesidad y los defectos cognitivos.
Bibliografía
Marshall WF, Nonaka S. 2006. Cilia: sintonizando la antena de la célula. Current biology. 16:R604-R614.
Satir P, Christensen ST. 2007. Overview of structure and function of mammalian cilia. Annual review of phisiology. 69:377-400.