Hay árboles genealógicos y poblaciones donde el principio de apareamiento panmítico se rompe esencialmente. Normalmente se trata de pequeñas comunidades relativamente aisladas geográfica o socialmente. En estas comunidades la endogamia, es decir el apareamiento consanguíneo de varios órdenes (un padre – un hijo, hermanos, primos, tío – sobrina, etc.), puede estar muy extendida.
El problema de la influencia de la endogamia en la población es el problema importante de la genética 1. Ahora este problema todavía no ha recibido la solución final.
En la mayoría de los casos la endogamia en un árbol genealógico tiene en cuenta sobre tres generaciones. Son los parientes más lejanos que se tienen en cuenta en el apareamiento consanguíneo hay primos segundos.
La endogamia se tiene en cuenta con la ayuda del factor de endogamia.
El factor de endogamia es la probabilidad de que dos alelos sean idénticos por origen.
Consideremos el método de cálculo del factor de endogamia en un árbol genealógico ofrecido por Wright 1, 2.
Para el hallazgo del factor de endogamia es necesario:
encontrar para los descendientes que se casan todos los caminos en un árbol genealógico que a través del antepasado común conectan estos descendientes.
contar número de pasos (alternancias de generaciones) en cada camino.
Entonces el factor de endogamia es igual:
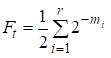 ……(1)
……(1)
donde r hay número de caminos, m1, m2, … mi- número de pasos en cada camino.
Una vez más observaremos que el número de sumandos en la suma (1) es igual al número de caminos que a través de los antepasados comunes conectan el matrimonio.
Consideremos casos concretos de árboles genealógicos, figura 1. En esta figura el cuadrado significa un individuo masculino el círculo – individuo femenino. Las líneas punteadas y las cifras idénticas especifican los caminos posibles en un árbol genealógico que a través del antepasado común (el hombre o la mujer) conectan de los descendientes el casamiento consanguíneo. En algunos sitios de los caminos, Figura 1с, hay dos dígitos 1 y 2. Especifica que tales sitios son el común para las varias maneras.
El matrimonio un padre – hija es incesto, Figura 1а. El descendiente (mujer) está conectado con el antepasado (hombre) allí 1 camino con un paso, es decir, m = 1. Por lo tanto, hay factor de endogamia.
Ft = 1/2.2-1 = 1/4
Hay matrimonio un hermano – hermana (hermanos), Figura 1b. Los dígitos especifican el número del camino. El número de dígitos idénticos en el camino del hombre que se casa con la mujer es igual al número de pasos.
Deje que cada descendiente que se ha casado conecta con su pareja dos caminos con dos pasos en cada camino, es decir, m = 2. Por lo tanto, hay factor de endogamia.
Ft = 1/2.(2-2 + 2-2) = 1/4
El matrimonio entre primos se muestra en la Figura 1с.
Figura 1. Algunos tipos de endogamia en un árbol genealógico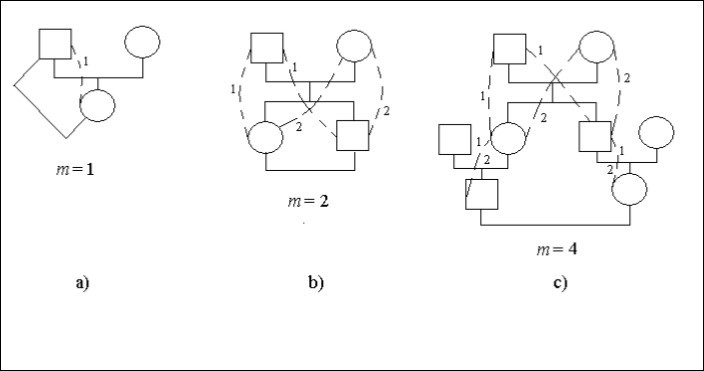
Cada descendiente que se ha casado conecta con pareja las 2 vías con 4 pasos en cada vía, es decir, m = 4. Por lo tanto, hay factor de endogamia.
Ft = 1/2.(2-4 + 2-4) = 1/16
La fórmula (1) es fácil de entender desde la siguiente lógica.
Obviamente hay 1/2 genes del hijo es el común con cada padre, 1/4 – con el abuelo y la abuela, 1/8 – con el bisabuelo y la bisabuela, etc. Si de la mujer a cualquier antepasado lleva el camino con η pasos la parte de los genes comunes con este antepasado es igual 2-η. Si desde el hombre hasta cualquier antepasado se sigue el camino con μ pasos, la parte de los genes comunes es igual a 2-μ. En el matrimonio del hombre y la mujer con el ancestro común la cuota de los genes comunes con este ancestro será 2-η. 2-μ = 2-m donde m = η + μ es el número total de pasos a través del ancestro común desde la mujer hasta el matrimonio del hombre. El tamaño 1/2 2-m muestra la probabilidad de que el gen elegido casualmente en la mujer sea idéntico por origen al gen elegido casualmente en el hombre. El mismo tamaño da la contribución al factor de endogamia Ft una vía a través del ancestro común desde la mujer hasta el hombre que se casa.
Por ejemplo, si el ancestro común estará lejos de casarse en η = μ = 5 generaciones la probabilidad de que tengan el gen común es igual. 1/2 2-(5+5) = 1/2048 Este tamaño en el análisis del apareamiento puede ser despreciado. (Tabla 1)
Tabla 1.Factores de consanguinidad para los algunos matrimonios consanguíneos se muestran además.
| El tipo de matrimonio consanguíneo matrimonio | Factor de consanguinidad Ft | Primos segundos | 1/32 |
| Tío primo – niece | 1/32 |
| Third cousins | 1/64 |
The inbreeding factor Ft is the probability of two alleles in posterity are identical by origin i.e. occur from same gene of ancestor 1. Or, that too most, the inbreeding factor is a probability of the allele origin in posterity is consanguineous. Hence, the probability of the non-consanguineous origins of identical alleles in homologous chromosomes is equal 1 – Ft.
Let’s find total probability of the recessive homozygote aa occurrence (eventаа) at random mating for an autosomal genes 3:
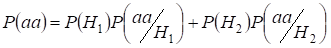 …….(2)
…….(2)
This formula follows from the following logic of mating: el origen de alelos idénticos en autosomas homólogos es no consanguíneo (la probabilidad de esta hipótesis es P(H1) = 1- Ft )también surge el homocigotoаа (la probabilidad condicional de este evento es P(aa/H1) = q2) o el origen de alelos idénticos es consanguíneo (la probabilidad de esta hipótesis es P(H2) = Ft ) y surge el homocigotoаа (la probabilidad condicional de este evento es P(aa/H2) = q). La probabilidad q de la aparición del homocigoto aa en este último caso viene determinada por que si el alelo a está en un autosoma entonces en el origen consanguíneo del homocigoto el mismo alelo debe estar necesariamente y en el autosoma homólogo.
Por lo tanto, la probabilidad total de ocurrencia del homocigoto recesivo es igual:
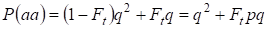 ……….(3)
……….(3)
Donde se tiene en cuenta p = 1 – q está la frecuencia del alelo dominante a.
De forma similar se encuentra la probabilidad total de ocurrencia del homocigoto dominante AA en el apareamiento aleatorio:
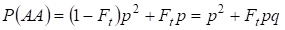 ……….. (4)
……….. (4)
Así, según (3) y (4) la frecuencia total de homocigotos (dominantes y recesivos) en el árbol genealógico endogámico aumenta en 2Ftpq. Por lo tanto, la frecuencia de los heterocigotos disminuye para el mismo tamaño y llegar a ser igual 2pq – 2pqFt = 2pq(1-Ft) ya que la suma de las frecuencias de todos los cigotos es igual a 1.
En vista del factor de endogamia y también (3) y (4) la ley de Hardy – Weinberg en la siguiente generación del árbol genealógico endogámico para un genoma autosómico necesita escribirse las distribuciones de los genotipos de los cigotos como 4:
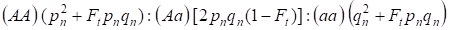 …………(5)
…………(5)
Donde pn es la frecuencia de alelos dominantes A en la generación n, qn – la frecuencia de alelos recesivos a, por lo que pn + qn = 1.
La relación (5) se refiere también a la relación de Wright 5. La relación de Wright muestra que en un árbol genealógico endogámico después del apareamiento consanguíneo de los individuos se observan los siguientes efectos: hay aumento de la fracción de homocigotos autosómicos en un árbol genealógico en Ft pnqny reducción de la fracción de heterocigotos autosómicos en (1-F1) en comparación con una población panmítica.
Pero ya a través de una generación se restablece el equilibrio Hardy – Weinberg que especifica la pequeña importancia del apareamiento consanguíneo separado para un árbol genealógico:
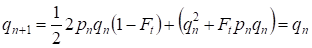 ………(6)
………(6)
Obviamente, para una población en su conjunto el acto separado del apareamiento consanguíneo tendrá aún menor importancia que para un árbol genealógico.
Se observa el mismo efecto de la variación de la frecuencia de los genotipos del cigoto y para los genes ligados al sexo.
La distribución de los genotipos ligados al cromosoma Х en el árbol genealógico panmítico se describe por la ley de Hardy – Weinberg como 1:
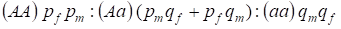 ………..(7)
………..(7)
La frecuencia de un alelo dominante A la designaremos en los hombres pm y en las mujeres pf. Para los alelos recesivosesa es en consecuencia qm y qf .
En el apareamiento hay una relación de genotipos en las mujeres según el producto (pf + qf) (pm +qm).
Hallemos la probabilidad total de ocurrencia del homocigoto recesivo aa en el apareamiento aleatorio para los genes de la hija ligados al cromosomaХ bajo la fórmula (2). La lógica de los acontecimientos es similar a la del caso de los autosomas, salvo que la probabilidad condicional de aparición de un genotipoаа a partir de los alelos no consanguíneos según (7) es igual P(aa/H1) = qmnqfn. Para los alelos consanguíneos, la probabilidad condicional de que aparezca un genotipoаа es igual a P(aa/H2) = qf(n+1). En las hijas, es decir, en la generación n+1 la probabilidad de ocurrencia de un homocigoto aa está determinada por que si el alelo a está en un cromosoma Х entonces en el origen consanguíneo del homocigoto el mismo alelo debe estar necesariamente y en el cromosoma Х homólogo.
Por lo tanto, la probabilidad total de ocurrencia del homocigoto recesivo es igual:
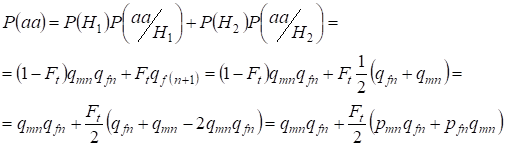 ……(8)
……(8)
donde pmn es la frecuencia de los alelos dominantes A en la generación n en los hombres y pfn – en las mujeres. Para los alelos recesivos a en consecuencia qmn y qfn . Además se utiliza qmn =1 – qmn y pfn = 1 – qfn.
De manera similar encontrar la probabilidad total de ocurrencia del homocigoto dominante con el uso:
 ,
,
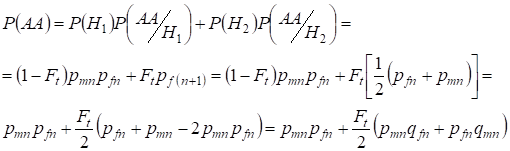 ……….(9)
……….(9)
y un heterocigoto P(Aa) = 1- P(aa) – P(AA) encontraremos la distribución de los genotipos en Х-cromosomas del árbol genealógico endogámico en el apareamiento:
 ……..(10)
……..(10)
Se demuestra simultáneamente que al igual que en los autosomas los aditivos endogámicos en las frecuencias de homocigotos dominantes y recesivos en los cromosomas ligados al sexo son idénticos.
Otra forma de hallar la relación (10) consistirá en lo siguiente.
Hallemos primero la probabilidad total de ocurrencia de un heterocigoto Aa, es decir, P(Aa) en la oportunidad del apareamiento consanguíneo al azar. Esta probabilidad se desprende de la siguiente lógica de apareamiento el origen de los alelos es no consanguíneo (la probabilidad de esta hipótesis es P(H1) = 1 – Ft ) también surge el heterocigoto Aа (la probabilidad condicional de este evento es P(Aa/H1) = pmnqfn + pfnqmn, véase la fórmula (7)) o el origen de los alelos es consanguíneo (la probabilidad de esta hipótesis es P(H2) = Ft) y surge el heterocigoto Aа (la probabilidad condicional de este evento es igual a cero P(Aa/H2) = 0 ). La última afirmación se define por el hecho de que en el origen consanguíneo los alelos similares en los cromosomas Х homólogos no pueden surgir el heterocigotoАа formado por alelos diferentes.
Por lo tanto, utilizando la fórmula de la probabilidad total encontramos la probabilidad total de la ocurrencia del heterocigoto Aa:
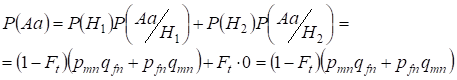 ……..(11)
……..(11)
La reducción de la frecuencia de los heterocigotos Aa se produce como se ha demostrado anteriormente debido al aumento idéntico de la frecuencia de los homocigotos dominantes AA y recesivos aa. Por ejemplo, el aumento de la frecuencia de homocigotos aa recesivos es igual:
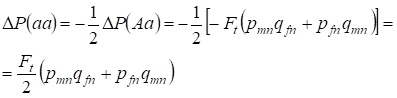 ……..(12)
……..(12)
donde la reducción de la frecuencia de heterocigotos Aa relativa al árbol genealógico panmítico según (7) y (11) es igual. ΔP(Aa) = – Ft (pmnqfn + pfnqmn) Utilizando (11) y (12) para actualizar (7) para los casos de endogamia encontramos la distribución de genotipos (10) en los cromosomas Х en el árbol familiar de las hijas endogámicas.
En la siguiente generación la frecuencia de los alelos recesivos a en las mujeres es igual:
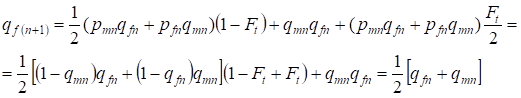 ……..(13)
……..(13)
Eso cumple plenamente con la misma frecuencia en ausencia de endogamia, es decir, hay una restauración del equilibrio Hardy – Weinberg o es más exacta la aproximación al equilibrio.
La relación (7) y (13) muestran en su conjunto las frecuencias alélicas en un árbol genealógico para la cuenta de la endogamia no varían. Sólo hay una redistribución de alelos. Una parte de los alelos dejan los heterocigotos y pasan a los homocigotos.