1. Cilium
2. Flagellum
3. Structuur
4. Beweging
5. Vorming
Microtubuli zijn cytoskeletonderdelen met belangrijke functies in de celfysiologie. De microtubuli scaffold van het cytoplasma is zeer plastisch dankzij de polymerisatie- en depolymerisatiecapaciteit van microtubuli. Niet alle celmicrotubuli zijn echter onderhevig aan deze krimp- of groeifase. Cilia, flagella en centriolen/basislichamen zijn celstructuren die zeer stabiele (aantal en lengte) en sterk georganiseerde microtubuli bevatten. Op deze pagina behandelen we cilia en flagella.
1. Cilium
Cilia zijn dunne en lange celuitsteeksels met een diameter van ongeveer 0,25 µm en een lengte van ongeveer 10 tot 15 µm, die voorkomen in dierlijke cellen en sommige eencellige eukaryotische soorten. Zij zijn gewoonlijk dicht opeengepakt aan het vrije oppervlak van epitheelcellen (figuren 1 en 2), zoals het epitheel van de ademhalingswegen, het epitheel van de voortplantingskanalen, de kieuwen van vissen en tweekleppigen, enz. Cilia zijn beweeglijke structuren en hun voornaamste functie is het verplaatsen van de omringende vloeistof, zoals het slijm van de luchtwegen, water rond het kieuwepitheel, maar ook de eicel in de vrouwelijke eileider. Veel eencellige organismen kunnen zich voortbewegen door trilhaartjes, en andere kunnen ze gebruiken voor het opwekken van waterwervelingen om voedsel te vangen. Embryo nodal cilia zijn betrokken bij het initiëren van de links-rechts as tijdens de embryonale ontwikkeling van gewervelde dieren. De beweging van cilia lijkt op kloppen, waardoor de vloeistof parallel aan het celoppervlak wordt gestimuleerd.
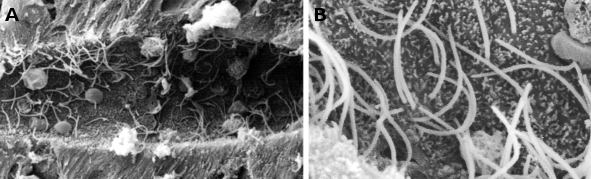
Figuur 1. Rasterelektronenmicroscopiebeelden van het centrale kanaal van het ruggenmerg van een lamprei. Veel cilia kunnen worden waargenomen (bij hogere vergroting in B) en kleine microvilli aan het apicale domein van ependimale cellen.
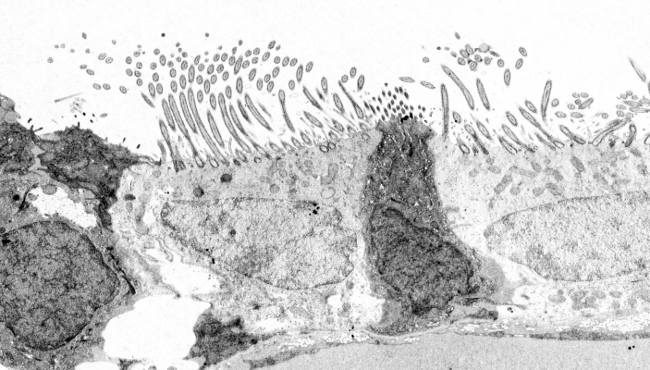
Figuur 2. Transmissie-elektronenmicroscopiebeelden van het respiratoire epitheel. Cellen met helder cytoplasma vertonen veel cilia in hun apicale oppervlak.
Er zijn cilia die niet kunnen bewegen, en dus niet bedoeld zijn voor vloeistofbeweging. Deze cilia staan bekend als primaire cilia. De meeste cellen die tot nu toe zijn bestudeerd (met uitzondering van rode bloedcellen) dragen primaire cilia: eileidercellen, neuronen, chondrocyten, ectodermcellen, mesenchymale cellen, urine-epitheelcellen, hepatocyten, en zelfs gekweekte cellen. Aanvankelijk werden primaire cilia beschouwd als niet-functionele trilharen. In het ciliaire membraan werden echter veel receptortypes en ionenkanalen aangetroffen, zodat ze als celsensorische structuren werden beschouwd. Zo worden olfactorische receptoren aangetroffen in cilia van hun dendrieten, en de uitwendige segmenten van staafjes en kegeltjes van het netvlies zijn eigenlijk gemodificeerde cilia. Sommige receptoren zitten hoger opeengepakt in de ciliaire membraan dan in andere plasmamembraandomeinen. Bovendien is er een grote verscheidenheid van moleculen in het inwendige van de cilia betrokken bij signaaltransductierollen. De hogere oppervlakte/volume-verhouding van een cilium maakt intraciliaire moleculaire reacties intenser en efficiënter dan wanneer deze zich buiten het cilium zouden bevinden. Naast chemische signalen kunnen primaire cilia ook vloeistofbewegingen buiten de cel waarnemen en als mechanoreceptoren werken.
2. Flagellum
Flagella lijken op cilia, maar ze zijn veel langer, ongeveer 150 µm lang, en iets dikker. Ze zijn veel minder talrijk dan trilharen in cellen. De belangrijkste functie van flagellen is het bewegen van de cel. De beweging van flagellen verschilt van die van cilia, omdat de bewegingsrichting loodrecht op het celoppervlak staat (niet evenwijdig), dat wil zeggen in de richting van de lengteas van het flagellum. Flagella kunnen vaak worden waargenomen in beweeglijke cellen zoals eencelligen en sperma.
Structuur
Cilia en flagella zijn complexe structuren die meer dan 250 verschillende eiwitten bevatten. Beide delen dezelfde centrale organisatie van microtubuli en andere geassocieerde eiwitten, samen bekend als axoneme, en begrensd door plasmamembraan (figuur 3). Naast axoneme zijn er vele oplosbare moleculen binnen de cilia/flagella die de matrix vormen. Het axonoom is opgebouwd uit 9 paren microtubuli rond een ander centraal paar microtubuli. Deze organisatie kan worden geschreven als (9 x 2) + 2. Bij primaire cilia ontbreekt het centrale paar microtubuli. Elke microtubulus van het centrale paar bestaat uit 13 protofilamenten, maar de microtubuli van de perifere paren delen enkele protofilamenten met elkaar. Zo wordt een perifeer paar gevormd door A- en B-microtubuli. De A-microtubulus bevat 13 protofilamenten en de B-microtubulus bevat 10 of 11 protofilamenten, waarvan er 2 of 3 worden gedeeld met de A-microtubulus.

Figuur 3. Belangrijkste moleculaire componenten van cilia en flagella. In primaire cilia is het centrale paar afwezig.
De microtubule-organisatie van het axoneem is het resultaat van een schavot van eiwitten. Twaalf eiwitten zijn reeds gevonden als bestanddelen van het axonoom die betrokken zijn bij het in stand houden van de microtubule organisatie. De naburige perifere microtubuleparen zijn onderling verbonden door nexine. In elk paar is de A-microtubule door eiwitspaken verbonden met een centrale ring die het centrale paar of de centrale microtubuli bevat. Dinein is een motoreiwit dat geassocieerd is met de perifere microtubuli die betrokken zijn bij de beweging van cilia en flagella.

Figuur 4. Ultrastructuur van een cilium van een ependimale cel van het ruggenmerg. (9+2)x2 betekent 9 perifere paren en 1 centraal paar microtubuli.
Microtubuli worden gepolymeriseerd vanuit basale lichaampjes (figuren 3 en 4). Het basaallichaam bestaat uit 9 triplet microtubuli die een cilinder vormen (vergelijkbaar met centriolen). Zij missen een centraal paar microtubuli, zodat het (9×3)+0 is. In elk triplet heeft slechts één microtubule (A-microtubule) een volledige set protofilamenten, terwijl B en C-microtubuli sommige daarvan met elkaar delen. Vanuit het basaallichaam groeien A- en B-microtubuli en vormen de perifere microtubuli van het axonoom. Net boven het basaallichaam bevindt zich een gebied van de cilia dat bekend staat als overgangszone met de 9 perifere paren en geen centraal paar. Onmiddellijk na de overgangszone bevindt zich de basale plaat, van waaruit het centrale paar microtubuli wordt gepolymeriseerd om het axoneem te voltooien. Alle microtubuli hebben het plus-einde naar de top van het cilium/flagellum. Het proximale uiteinde van het basale lichaam (het binnenste, of minus uiteinde van de microtubuli) is verankerd aan het celcytoskelet door middel van lange eiwitvezels die ciliaire wortelknopen worden genoemd
Naast het axonem hebben cilia/flagella nog andere compartimenten. Het membraan bevat veel receptoren en kanalen om de omgeving waar te nemen, vooral in primaire cilia. De vloeibare fase van het inwendige wordt matrix genoemd, die helpt bij het georganiseerd houden van de hele structuur en verantwoordelijk is voor het doorgeven van de informatie die door de membraanreceptoren is verzameld. Andere afzonderlijke gebieden zijn het basale lichaam aan de basis en het apicale deel van het cilium/flagellum, dat eiwitten bevat die de pluspunten van de microbolletjes stabiliseren.
4. Beweging
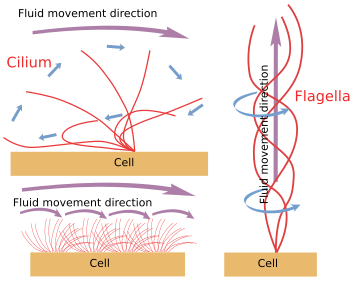
Figuur 6. Modellen voor cilium- en flagelabeweging. Ze genereren verschillende vloeistofbewegingsrichtingen.
Wanneer cilium/flagellum mechanisch van de cel worden losgemaakt, blijven ze bewegen tot de ATP-voorraad is uitgeput. Dit betekent dat het bewegingsmechanisme intrinsiek is (figuur 6). In feite ontstaat de beweging door het schuiven van een perifeer paar microtubuli over de buurman. Nexine en spoke-eiwitten verhinderen de desorganisatie van het axonoom, maar maken deze bewegingen mogelijk. Dinein is het motoreiwit dat verantwoordelijk is voor de schuifbeweging. De energie wordt geleverd door ATP. Dineïnen worden met hun bolvormige deel verankerd aan de A-microtubule van één perifeer paar en met hun staartdeel aan de B-microtubule van het aangrenzende paar. Het moleculaire mechanisme is vergelijkbaar met dat van de cytosolische dineïnen, maar in plaats van een lading te vervoeren, wordt een microtubule in beweging gebracht. Voor een efficiënte beweging is coördinatie van de dineinen van het axonoom nodig. Calciumgolven in het inwendige van de cilia/flagella kunnen de activering van dineïnen coördineren en de bewegingsfrequentie wijzigen wanneer dat nodig is. Het is van belang op te merken dat niet alle dineinen tegelijkertijd geactiveerd hoeven te worden, maar synchroon.
5. Vorming
Tijdens de differentiatie produceren de cellen alle noodzakelijke cilia en flagella voor hun normale fysiologie. Dit betekent dat ze allemaal vanaf nul moeten worden gegenereerd. Microtubuli van axonemen worden gekernd uit A- en B-microtubuli van de basale lichamen, dus per cilium/flagellum is één basaal lichaam nodig. Hoe worden meerdere basale lichamen gevormd? Er zijn ten minste drie manieren om basa-lichaampjes te produceren: a) door centriolen te gebruiken als sjabloon voor de nucleatie van basa-lichaampjes; b) uit amorf materiaal dat bekend staat als deuterosoom; c) in planten zijn er verschillende eiwitaggregaten die basa-lichaampjes kunnen nucleeren.
Veel menselijke pathologieën zijn het gevolg van cilium/flagella defecten. Ze staan bekend als ciliopathieën, en omvatten willekeurige lateraliteit, verkeerde sluiting van de neurale buis, polydactylie, cystische nier-, lever- en pancreaspathologieën, netvliesdegeneratie, obesitas, en cognitieve defecten.
Bibliografie
Marshall WF, Nonaka S. 2006. Cilia: afstemmen op de antenne van de cel. Huidige biologie. 16:R604-R614.
Satir P, Christensen ST. 2007. Overview of structure and function of mammalian cilia. Annual review of phisiology. 69:377-400.