Introduction
Twee visuele signalen die gelijktijdig verschijnen worden sneller waargenomen dan een van beide signalen alleen (Hershenson, 1962; Raab, 1962). Een deel van dit redundante target effect (RTE) kan stochastisch worden verklaard door een ‘horse-race’ model. Dat wil zeggen, als twee stimuli worden verwerkt in parallelle en onafhankelijke kanalen waarvan de verwerkingssnelheid willekeurig varieert van trial tot trial, dan wint bij elke presentatie de snelste stimulus de race en bereikt de detectiedrempel om de respons te triggeren; daarom is het gemiddeld waarschijnlijker dat twee stimuli een snellere respons opleveren dan de gemiddelde responstijd op één stimulus die in een van de twee kanalen wordt verwerkt.
Analyses van cumulatieve frequentieverdelingen hebben echter aangetoond dat er een extra bijdrage is aan de RTE die niet kan worden verklaard door een racepaardenmodel tussen twee onafhankelijke kanalen. Miller (1982) stelde een “co-activatie” model voor waarin de twee signalen worden opgeteld in een activeringspool. Miniussi e.a. (1998) toonden aan dat redundante doelen kortere latenties veroorzaakten voor P1 en N1 gebeurtenisgerelateerde hersenpotentialen, hetgeen erop wijst dat neurale sommatie vroeg in de visuele pathway optreedt. Experimenten in hemianopische patiënten hebben ook aangetoond dat de RTE niet volledig kan worden verklaard door een paardenrace om de waarnemingsdrempel te bereiken, aangezien een RTE zelfs manifest kan zijn in de afwezigheid van fenomenaal bewustzijn van een van de twee stimuli (Marzi et al., 1986; Tomaiuolo, 1997; de Gelder et al., 2001; Leh et al., 2006a,b). Inderdaad, in hemispherectomiepatiënten genereert een redundant doelwit in het blinde veld niet alleen een RTE, de RTE wordt vergroot wanneer de redundante stimulus een gestaltpatroon completeert (Georgy et al., 2016). Deze observaties in hemianopics suggereren dat neurale summatie subcorticaal plaatsvindt en niet afhankelijk is van primaire visuele cortex.
Het cumulatieve indirecte bewijs heeft de superieure colliculus (SC) geïmpliceerd als het substraat voor de neurale summatie die bijdraagt aan de RTE. Direct bewijs is echter nog niet gerapporteerd. Het huidige onderzoek bestudeert het neurale pad dat de visuele signalen doorgeeft die geïntegreerd worden in de SC. Het SC bestaat uit oppervlakkige en diepe lagen. De diepe lagen van de colliculus ontvangen afferenten, zowel direct als via de basale ganglia, van de oculomotor cortex (Hikosaka en Wurtz, 1983; Pare en Wurtz, 1997; Sommer en Wurtz, 2000). Visuele afferenten naar de oppervlakkige lagen van de colliculus worden echter zowel vanuit het netvlies (via het retino-tectale kanaal) als vanuit de vroege visuele cortex (Fries, 1984) doorgegeven, via het brachium van het SC.
Omdat neurale summatie effecten zijn aangetoond bij neurologische patiënten die geen visuele cortex hebben, zouden de visuele signalen die in de colliculus worden samengevat rechtstreeks vanuit het netvlies naar de oppervlakkige lagen van de colliculus kunnen worden doorgegeven via het retino-tectale kanaal. Als dit het geval is, zou neurale sommatie kunnen worden opgeheven door laesies van het brachium van het SC.
Lesies die het brachium van het SC verstoren zijn zeer zeldzaam. Hier rapporteren wij individuele casestudies van twee patiënten met unilaterale subcorticale laesies die de SC uitschakelden van visuele input, en die geen RTE manifesteerden. Eén patiënt had een laesie die zowel de pulvinar nucleus van de thalamus, de rostrale SC, als het brachium beschadigde. Bij de andere patiënt, met schade aan de dorsale middenhersenen als gevolg van een traumatisch hersenletsel, bleek het SC intact te zijn, maar het brachium ervan was verstoord.
Materialen en Methoden
Deelnemers
Twee neurologische patiënten, elk met haar eigen leeftijdsgecorrigeerde controlegroep, werden getest in een experiment om de RTE te meten.
RE was 71 jaar oud op het moment van testen. Een posterieure thalamische bloeding 3 jaar eerder vernietigde de mediale pulvinar, ventro-laterale thalamische kernen en de achterste ledemaat van het interne kapsel met schade uit te breiden tot de dorsale middenhersenen met inbegrip van het pretectum en rostrale SC (figuur 1) Probabilistische DTI tractografie (Behrens et al., 2003, Behrens et al., 2007) bevestigd dat de laesie het brachium van het SC (figuur 2) vernietigd. RE was verlamd aan de linkerzijde van haar lichaam met verlies van gevoel in het linker gezicht en arm. Ze had abnormale oogbewegingen, waaronder ooglidretractie, verlamde verticale oogbewegingen en verminderde convergentie, die een gevolg waren van de uitbreiding van de laesie in de dorsale middenhersenen.
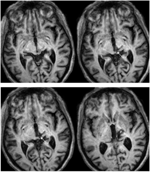
FIGUUR 1. Hoge resolutie (0,7 mm3) T1-gewogen MRI van patiënt RE. Axiale doorsneden van ventraal (linksboven) naar dorsaal (rechtsonder) tonen de laesie in de rechter dorsale middenhersenen en thalamus. Uitbreiding van de laesie in de rostrale superieure colliculus (SC) getoond door witte pijlen.
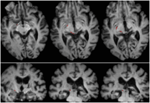
FIGUUR 2. Tractografie van de retinotectale tractus bij patiënt RE, in co-registratie met T1-gewogen axiale doorsneden (boven van ventraal naar dorsaal) en coronale doorsneden (onder van anterieur naar posterieur). Het verloop van de retinotectale tractus wordt getoond in de linker hemisfeer (rood). Geen stroomlijn werd getraceerd in de rechter hersenhelft en, door vergelijking met de intacte hemisfeer kan worden gezien dat de laesie vernietigd het brachium van de SC in de rechter hersenhelft. De stroomlijn in het rood werd gegenereerd met probabilistische tractografie met behulp van FSL FDT (FMRIB Diffusion Toolbox; Behrens et al., 2003, Behrens et al., 2007; http://fsl.fmrib.ox.ac.uk/fsl/fslwiki). Diffusie-gewogen echoplanaire magnetische resonantie beelden werden verkregen met een resolutie van 1,5 mm3 met 32 isotropisch verdeelde diffusie-coderende richtingen (b = 800) en een basislijn (b = 0). Herhalingstijd = 2 s, en echotijd = 35 ms. Voor de probabilistische tractografie, markeerden we handmatig de start regio (dat wil zeggen, trok zaad maskers) op de optische tractus van elk halfrond net achter het chiasme en een doel regio (doelmasker) op de SC.
ML was 23 jaar oud op het moment van testen. Ze had een ernstig traumatisch hersenletsel opgelopen bij een verkeersongeval 7 jaar eerder, resulterend in diffuus axonaal letsel met bloeding in het rechter putamen en dorsale middenhersenen. Details van haar voorgeschiedenis, neurologisch onderzoek, en neuroimaging bevindingen zijn gerapporteerd door (Poliva et al., 2015). Hoewel links was enige motorische beperkingen, waaronder spasticiteit en ataxie, had ze herwonnen onafhankelijkheid. Haar belangrijkste handicap was ernstige auditieve agnosie als gevolg van schade aan de hersenstam auditieve paden met inbegrip van de linker inferior colliculus. Haar gezichtsscherpte en gezichtsvelden waren niet aangetast en er was geen visuele verwaarlozing of extinctie. Oculomotorische tekenen van middenhersendisfunctie waren onder andere macro-square wave jerks en convergentiespasme bij verticaal staren (meer naar beneden dan naar boven.) Er was geen ptosis of pupilafwijking. Hoge resolutie MRI toonde een cystische holte in het rechter putamen op de plaats van haar vorige bloeding, en een kleine periventriculaire laesie op het rechter onderste pons, in de regio van de inferieure cerebellaire pedunkel (figuur 3). Ze had bijna volledige avulsie van de linker colliculus inferior, waarbij alleen de meest mediale en caudale delen werden gespaard. De schade strekte zich uit tot in het laterale pretectum en het midbrain tegmentum, waarbij de brachia van de superieure en inferieure colliculi werden vernietigd, en ventraal tot in de rode nucleus. Probabilistische DTI-tractografie bevestigde dat de laesie het brachium van het SC verwoestte (figuur 4).
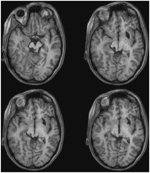
FIGUUR 3. T1-gewogen MRI met hoge resolutie (0,7 mm3) van patiënt ML. Axiale doorsneden van ventraal (linksboven) naar dorsaal (rechtsonder) tonen schade aan de linker colliculus inferior en het mesencefalisch tegmentum ventraal en lateraal van het SC.
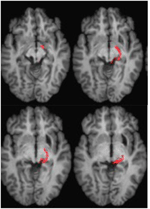
FIGUUR 4. DTI tractografie bij patiënt ML toont het netvlies-tectale kanaal in de rechter hemisfeer (rood), gecoördineerd met T1-gewogen MR-beelden in het axiale vlak van ventraal (linksboven) naar dorsaal (rechtsonder). Diffusie-gewogen echo-planaire magnetische resonantie beelden werden verkregen bij 2 mm3 resolutie met 63 isotropisch verdeelde diffusie-coderende richtingen (b = 1000) en een basislijn (b = 0). Herhalingstijd = 2 s, en echotijd = 35 ms. Tractografie werd uitgevoerd zoals beschreven in figuur 2.
Neurologisch gezonde controle proefpersoon voor patiënt RE omvatte 11 oudere volwassenen (zes vrouwen en vijf mannen, gemiddelde leeftijd: 68,5, range 59-73). Zij werden gerekruteerd uit het deelnemerspanel van de Bangor Universiteit. De deelnemers hadden geen bekende neurologische, psychologische, psychiatrische of cognitieve beperkingen en alle deelnemers hadden een normaal/gecorrigeerd tot normaal gezichtsvermogen.
De neurologisch gezonde controlepersonen voor patiënt ML bestonden uit 11 studenten en postdoctorale studenten van Bangor University, variërend in leeftijd van 18-26 jaar (zeven vrouwen en vier mannen).
Apparatuur en kijkomstandigheden
Presentation® programmeersoftware op een PC computer registreerde reactietijd (RT) responsen en genereerde stimuli die werden gepresenteerd op een Dell monitor (12.5″ × 25″, verversingssnelheid 60 Hz.) geplaatst op ooghoogte 57 cm voor de deelnemers met binoculair kijken in gedimd licht. Een kin- en hoofdsteun werden gebruikt om de stabiliteit van het hoofd te garanderen.
Stimuli
De experimentele stimuli bestonden uit ongevulde witte markervierkanten (1 cm × 1 cm) gepositioneerd 8° links en rechts van een kleine fixatie box (0.2 cm × 0.2 cm) in het midden van het scherm. Doelstimuli bestonden uit het invullen van een of beide perifere markervakjes om een effen wit vierkant te produceren. Alle stimuli waren wit gepresenteerd op een grijze achtergrond.
Figuur 5 toont het verloop van een enkele proef. De marker vakken zichtbaar bleef gedurende het experiment. Na een inter-trial interval van 1750 ms, verscheen de fixatie box om de trial te starten. Na een willekeurig interval van 250 tot 750 ms (in stappen van 25 ms), verscheen een doelwit dat een eenvoudige druk op de spatiebalk van het toetsenbord vereiste, op de plaats van een of beide markeringsvakjes. Het doelwit was een gevuld wit vierkant, gegenereerd door het invullen van een of beide markeringsvakjes. De doelen verschenen willekeurig en met gelijke waarschijnlijkheid links, rechts, of beide tegelijk. Bij de vangstproeven (10% van de proeven) verscheen geen doelwit.
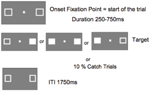
FIGUUR 5. ITI, inter-trial interval tussen de reactie van de deelnemer en het begin van het fixatiepunt waarmee de volgende test begint)
Procedure
Elke deelnemer werd in één sessie getest. Na het lezen van de instructies op het scherm, kregen de deelnemers voorbeelden te zien van alle stimuli die in de taak werden gepresenteerd (het fixatievak en elk van de drie doelpresentaties) en werd gevraagd om de herkenning van elk doel te bevestigen door op de spatiebalk te drukken. De instructies waren om de fixatie op het centrale vak te handhaven voor de duur van elke proef, en om zo snel mogelijk te reageren door het indrukken van de spatiebalk met de rechterhand zodra een doel verscheen.
Als de deelnemer verbaal en door het indrukken van de toets bevestigde dat alle instructies waren begrepen, werd overgegaan tot het presenteren van oefenproeven (10 van elke doelpresentatie conditie en 3 doelwit-afwezige ‘vang’ proeven). Tijdens het voltooien van de oefenproeven werd feedback op het scherm gegeven, waaronder de volgende mededelingen: (1) “Correct”, (2) “Probeer sneller te reageren!” en (3) “Reageer alleen als je een doel ziet.” Zodra de oefensessie was voltooid en zowel de deelnemer als de experimentator ervan overtuigd waren dat de taakinstructies correct waren begrepen, werd de deelnemer uitgenodigd om met het experiment te beginnen. De eerste 30 trials van elk experimenteel blok werden uitgesloten als oefen trials. Elk blok bestond uit een totaal van 233 trials: 70 rechts, 70 links, 70 beide, en 23 ‘catch’ trials. Halverwege het blok werd een korte pauze ingelast. Patiënt ML en de jongere controles werden in één blok getest. Vanwege de variabiliteit die samenhangt met de leeftijd en conditie van patiënte RE, werd zij getest in twee blokken op verschillende dagen; en haar leeftijdsgenootjes in de controlegroep werden ook getest in twee sessies op één dag.
Analyses
Voor elke controlegroep werd de mediane RT voor elke deelnemer in elke conditie (linkerdoelwit, rechterdoelwit, bilaterale doelen) berekend na uitsluiting van proeven die volgden op vangproeven en van proeven met RT’s van <100 ms of >800 ms. Een gepaarde t-test werd uitgevoerd om te bevestigen dat er geen asymmetrie was in de respons op unilaterale doelen rechts en links, en een gemiddelde RT voor unilaterale doelen werd berekend. RTE werd berekend door RTs voor bilaterale trials af te trekken van RTs voor unilaterale trials. Een gepaarde sample t-test vergeleek unilateraal met bilateraal om vast te stellen of de RTE statistisch betrouwbaar was.
Voor elke patiënt, na uitsluiting van trials volgend op vangst trials en die met RTs van <100 ms of >800 ms, vergeleek een gepaarde sample t-test de mediane RTs voor contralaterale en ipsilaterale unilaterale doelen. Zoals hieronder gerapporteerd, waren de RTs voor reacties op contralaterale doelen langer dan voor ipsilesionale unilaterale doelen. Om te bepalen of de aanwezigheid van een contralesionaal doel een statistisch betrouwbare RTE opleverde, vergeleek een gepaarde samples t-test de mediane RTs voor bilaterale doelproeven met unilaterale ipsilesionale doelproeven.
Om te testen of de RTE betrouwbaar kleiner was voor elke patiënt dan voor hun respectieve controlegroep, werd de RTE berekend voor elke controledeelnemer en werden de boven- en ondergrenzen van de 95%-betrouwbaarheidsintervallen (CI) berekend. De Z-score voor de RTE van elke patiënt werd berekend ten opzichte van hun controlegroep en getest op statistische betrouwbaarheid met de Crawford’s t-test.
P waarden voor gepaarde steekproef t-tests werden gerapporteerd voor twee staarten en voor Crawford t, één staart.
Resultaten
Patiënt RE
Controle Deelnemers: Het gemiddelde van de mediane RTs verschilde niet voor linker- en rechterantwoorden (t = 0,6.) Figuur 6 laat zien dat RTs korter waren (de RTE) voor bilaterale doelen dan voor ofwel linker (t = 6,8, p < 0,001, of rechter (t1,10) = 3,8, p = 0,003) unilaterale doelen. De mediane RT voor bilaterale doelpogingen werd afgetrokken van het gemiddelde van de mediane RT voor unilaterale doelen om een gemiddelde RTE te berekenen voor elke controle deelnemer. De gemiddelde RTE voor de controlegroep was 24,3 ms (SEM = 2,75).

FIGUUR 6. Mediane RTs voor patiënt RE, en gemiddelde van de mediane RTs voor deelnemers aan de controlegroep voor doelen die verschijnen in het linker gezichtsveld (‘Links’), rechter gezichtsveld (‘Rechts’), en tegelijkertijd in beide gezichtsvelden (‘Beide’).
Patiënt RE’s mediane RTs waren langer dan controles (figuur 6), en lagen boven de bovengrens van de 95% CI van de controlegroep gemiddelde RTs voor alle drie de condities (links = 457 ms; rechts = 449 ms; beide = 426 ms). De RTs voor unilaterale doelen in het contralesionale (linker) veld waren langer dan voor unilaterale doelen in het ipsilesionale (rechter) veld (t = 6.0, p < 0.001).
Belangrijker is dat er geen RTE was: de mediane RT voor bilaterale doelen (500 ms) was niet korter dan voor unilaterale doelen in het ipsilesionale (rechter) gezichtsveld (494 ms; t = 1,141), p = ns (figuur 6). Tabel 1 (boven) laat zien dat de RTE berekend voor patiënt RE significant minder was dan voor de RTE voor haar controlegroep.

TABLE 1. Redundant target effect voor patiënt RE (boven) en ML (onder), en gemiddelde RTE voor hun respectieve controlegroepen met 95%-betrouwbaarheidsintervallen (CI), Z-scores en Crawford t-test die elke patiënt vergelijkt met haar respectieve controlegroep.
Patiënt ML
Controle Deelnemers: Het gemiddelde van de mediane RTs verschilde niet voor responsen op linker en rechter doelen (t = 0,86. Figuur 7 laat zien dat de RTs korter waren (de RTE) voor bilaterale doelen dan voor ofwel linker (t = 4,7, p = 0,001, of rechter (t1,11) = 7,8, p < 0,001) unilaterale doelen.

FIGUUR 7. Mediane RTs voor Patiënt ML, en gemiddelde van de mediane RTs voor deelnemers aan de controlegroep voor doelen die verschijnen in het linker gezichtsveld, rechter gezichtsveld, en tegelijkertijd in beide gezichtsvelden.
De mediane RT voor bilaterale doelpogingen werd afgetrokken van het gemiddelde van de mediane RT voor unilaterale doelen links en rechts om een RTE te berekenen voor elke controle deelnemer.
De groepsgemiddelde RTE was 33 ms (SEM = 6,65).
De mediane RTs van patiënt ML waren langer dan die van de controles (figuur 7), en lagen boven de bovengrens van de 95% CI van de gemiddelde RTs van de controlegroep voor alle drie de condities (links = 322 ms; rechts = 307 ms; beide = 277 ms). De RTs voor unilaterale doelen in het rechter (contralesionale) veld waren langer dan voor unilaterale doelen in het linker (ipsilesionale) veld (t = 4,9, p = 0,001). Er was geen RTE. RTs voor bilaterale doelen (361 ms) waren niet korter dan voor unilaterale doelen in het ipsilesionale (linker) gezichtsveld (363 ms). Tabel 1 (onder) laat zien dat de RTE berekend voor patiënte ML significant minder was dan de RTE voor haar controlegroep.
Discussie
Eenvoudige RTs werden gemeten voor het detecteren van enkelvoudige doelen in het contralesionale of ipsilesionale veld, en van bilaterale doelen, bij twee patiënten bij wie het brachium van het SC beschadigd was in één hemisfeer. Bij één van de patiënten (een jonge vrouw) was de beschadiging in de linker hemisfeer het gevolg van traumatisch hersenletsel; en, hoewel de laesie de colliculus inferior betrof en zich uitstrekte tot in het tegmentum midbrain, was de beschadiging van het SC als zodanig niet duidelijk met hoge-resolutie neuroimaging. Bij de andere patiënt (een oudere vrouw) bevond de schade zich in de rechter hemisfeer en werd veroorzaakt door een hypertensieve bloeding die ook uitgebreide schade veroorzaakte aan de mediale pulvinar en laterale thalamus en zich uitstrekte tot in de dorsale middenhersenen met inbegrip van de rostrale SC. Bij beide patiënten was de eenvoudige RT om visuele doelen in het contralesionale veld te detecteren vertraagd in vergelijking met het ipsilesionale veld. Geen van beide patiënten vertoonde een ruimtelijk sommatie effect (RTE) voor doelwit detectie wanneer stimuli werden gepresenteerd in beide visuele velden.
Deze observaties werpen licht op de rol van het SC in doelwit detectie en, meer specifiek, de rol van visuele afferenten naar het SC. Posner et al. (1980) definieerden detectie operationeel als het vermogen om een arbitraire respons te geven op een visueel signaal (b.v. een simpele toetsdruk met een vinger, zoals in het huidige experiment). Zij stelden dat detectie een toewijzing van aandacht vereist om het doel te selecteren voor verwerking in een systeem met beperkte capaciteit dat prioriteit geeft aan het doel om naar te handelen.
Song et al. (2011) toonden aan dat inactivering van SC bij apen opvallende doelselectietekorten veroorzaakt die niet gemakkelijk kunnen worden verklaard als een eenvoudige stoornis in visuele perceptie of motorische uitvoering, en suggereerden dat het bijdraagt aan een meer algemene doelprioriteitenkaart. In dat experiment kregen apen twee stimuli in tegenovergestelde gezichtsvelden te zien en moesten ze met een reikrespons aangeven welke als eerste verscheen. Eén aap werd getraind om te reiken naar de stimulus die als eerste verscheen; en één aap werd getraind om te reiken naar het doelwit dat als tweede verscheen. In beide gevallen was er een sterke bias tegen het selecteren van het doelwit dat zich in het gezichtsveld contralateraal van de laesie bevond. Aangezien dezelfde bias werd gezien bij de aap die reikte naar het tweede doel dat verscheen, kon de bias niet worden toegeschreven aan een vertraging in het waarnemen van het contralesionale doel. Colliculaire inactivatie veroorzaakte geen stoornis in de perceptuele beoordeling van welke stimulus eerst verscheen. Integendeel, stimuli in het visuele veld contralateraal aan de geïnactiveerde colliculus werden benadeeld in het krijgen van prioriteit voor actie.
Zhaoping (2016) heeft recentelijk gewezen op een onderscheid tussen neurale saliëntiekaarten en prioriteitskaarten, en betoogd voor een evolutionaire migratie van een perceptuele saliëntiekaart van het optisch tectum/SC naar de primaire visuele cortex. Zij stelde voor dat saliëntiesignalen kunnen worden doorgegeven aan een prioriteitskaart in het SC zonder de eigenschap van feature tuning te projecteren.
Het brachium van het SC geeft afferente visuele signalen door aan de oppervlakkige lagen van de colliculus vanuit het netvlies via het retino-tectale tractus; maar de meerderheid van de visuele afferente vezels die door het brachium worden doorgegeven zijn projecties van de primaire visuele cortex. Indien het brachium verstoord is, kunnen signalen van de primaire visuele cortex naar de colliculus worden doorgegeven via de frontale oogvelden. Maar in de afwezigheid van directe projecties naar de oppervlakkige lagen van de colliculus vanuit het netvlies of vanuit de visuele cortex, zoals wij veronderstellen dat het geval is bij de patiënten met brachium laesies, duurt het langer voordat de colliculaire prioriteitskaart wordt geactiveerd, wat resulteert in tragere reacties op contralesionele visuele signalen.
Zoals in de inleiding opgemerkt, is de bijdrage aan de RTE die gebaseerd is op een probabilistisch race horse model afhankelijk van detectie van beide doelen; en van een race waarin de efficiëntie van onafhankelijke kanalen waarin de twee doelen worden doorgegeven gelijkwaardig zijn, zodat de uitkomst van de race om de detectiedrempel te bereiken willekeurig is van trial tot trial. Aangezien de detectiedrempel voor contralesionale signalen hoger is bij de hier gerapporteerde patiënten, kunnen redundante doelen niet profiteren op basis van een stochastische paardenrace; dat wil zeggen, de uitkomst van de race tussen ipsilesionale en contralesionale signalen is niet willekeurig – contralesionale signalen hebben meer kans om de race te verliezen dan te winnen.
Niettemin heeft onderzoek bij hemianopische patiënten aangetoond dat er ook een neurale sommatie component is die bijdraagt aan de RTE die niet afhankelijk is van detectie van beide doelen (Marzi et al., 1986; Tomaiuolo, 1997; de Gelder et al., 2001). Omdat commissurotomized patiënten een RTE blijken te hebben (Marzi et al., 1986; Reuter-Lorenz et al., 1995), is beargumenteerd dat interhemisferische integratie van signalen over de verticale meridiaan subcorticaal moet plaatsvinden.
Savazzi en Marzi (2004) toonden aan dat neurale sommatie niet optrad bij korte golflengte chromatische doelstimuli bij zowel neurologisch intacte mensen als patiënten met gespleten hersenen. Leh et al. (2006b) rapporteerden dat, terwijl een RTE optrad met achromatische stimuli in sommige hemianopische hemispherectomized patiënten, er geen RTE was in deze patiënten wanneer korte golflengte chromatische stimuli werden gebruikt als doelen. Korte golflengte (d.w.z., paars) stimuli activeren alleen S-kegeltjes in het netvlies. Retinale ganglion cellen die input ontvangen van S-cones projecteren niet direct naar het SC, noch naar magnocellulaire lagen van de laterale geniculate nucleus (De Monasterio, 1978). Aangezien projecties van de primaire visuele cortex naar het SC via het brachium van het SC alleen visuele signalen doorgeven van magnocellulaire geniculostriate afferenten (Schiller et al., 1979), suggereert het falen van S-cone stimuli om een RTE op te wekken dat neurale sommatie optreedt in het SC en afhankelijk is van visuele afferenten die doorgegeven worden via zijn brachium.
Er is dus convergerende evidentie in gespleten hersenen patiënten, hemianope patiënten met blindziendheid, en uit experimenten met korte golflengte stimuli, dat neurale sommatie optreedt in het SC. Niettemin, terwijl korte golflengte stimuli geen retinale ganglioncellen activeren die via het brachium naar het SC projecteren, kan niet worden geconcludeerd dat de colliculus volledig blind is voor dergelijke stimuli. Single unit opnamen in de SC van apen hebben aangetoond dat korte golfstimuli reacties activeren in de oppervlakkige lagen van de colliculus. Bovendien heeft tractografie aangetoond, in die hemisfeergectomiseerde patiënten met een RTE (d.w.z. blindziendheid), maar niet in die patiënten die geen aanwijzingen van blindziendheid vertoonden, dat de SC verbinding had met de intacte hemisfeer (Leh et al., 2006a).
De bevindingen van het huidige onderzoek leveren dus het eerste directe bewijs dat ruimtelijke sommatie optreedt in het SC dat afhankelijk is van transmissie van visuele signalen via zijn brachium.
Ethics Statement
Dit onderzoek werd uitgevoerd in overeenstemming met de richtlijnen en met goedkeuring van het protocol van de NHS Ethics Committee, Bangor, UK, en de School of Psychology Ethics Committee van Bangor University, UK. Mondelinge en schriftelijke geïnformeerde toestemming werd verkregen van alle deelnemers in overeenstemming met de Verklaring van Helsinki.
Author Contributions
MvK: bedacht het onderzoek, ontwierp en programmeerde het experiment, verzamelde en analyseerde patiëntgegevens en schreef mee aan het manuscript. KK: verzamelde controlegegevens, beoordeelde relevante literatuur, analyseerde controlegegevens en schreef mee aan het manuscript. RR: Rekruteerde patiënten en analyseerde neuroimaging studies inclusief virtuele dissectie met DTI tractografie, bereidde patiënt case histories voor en schreef mee aan het manuscript.
Conflict of Interest Statement
De auteurs verklaren dat het onderzoek werd uitgevoerd in afwezigheid van enige commerciële of financiële relaties die zouden kunnen worden opgevat als een potentieel belangenconflict.
Behrens, T., Woolrich, M. W., Jenkinson, M., Johansen-Berg, H., Nunes, R. G., Clare, S., et al. (2003). Karakterisering en voortplanting van onzekerheid in diffusie-gewogen MR beeldvorming. Magn. Reson. Med. 50, 1077-1088. doi: 10.1002/mrm.10609
PubMed Abstract | Google Scholar | Volledige Tekst | Google Scholar
Behrens, T. E., Berg, H. J., Jbabdi, S., Rushworth, M. F., and Woolrich, M. W. (2007). Probabilistic diffusion tractography with multiple fibre orientations: what can we gain? Neuroimage 34, 144–155.
Google Scholar
de Gelder, B., Pourtois, G., van Raamsdonk, M., Vroomen, J., and Weiskrantz, L. (2001). Unseen stimuli modulate conscious visual experience: evidence from interhemispheric summation. Neuroreport 12, 385–391. doi: 10.1097/00001756-200102120-00040
CrossRef Full Text | Google Scholar
De Monasterio, F. M. (1978). Properties of ganglion cells with atypical receptive-field organization in retina of macaques. J. Neurophysiol. 41, 1435–1449.
PubMed Abstract | Google Scholar
Fries, W. (1984). Cortical projections to the superior colliculus in the macaque monkey: a retrograde study using horseradish peroxidase. J. Comp. Neurol. 230, 55–76. doi: 10.1002/cne.902300106
PubMed Abstract | CrossRef Full Text | Google Scholar
Georgy, L., Celeghin, A., Marzi, C. A., Tamietto, M., and Ptito, A. (2016). De superieure colliculus is gevoelig voor gestalt-achtige stimulusconfiguratie in hemispherectomiepatiënten. Cortex 81, 151-161. doi: 10.1016/j.cortex.2016.04.018
PubMed Abstract | CrossRef Full Text | Google Scholar
Hershenson, M. (1962). Reaction time as a measure of intersensory facilitation. J. Exp. Psychol. 63, 289-293. doi: 10.1037/h0039516
CrossRef Full Text | Google Scholar
Hikosaka, O., and Wurtz, R. (1983). Visuele en oculomotorische functies van de apen substantia nigra pars reticulata. III. geheugen contingente visuele en saccade responsen. J. Neurophysiol. 49, 1268-1284.
PubMed Abstract | Google Scholar
Leh, S. E., Johansen-Berg, H., and Ptito, A. (2006a). Unconscious vision: new insights into the neuronal correlate of blindsight using diffusion tractography. Brain 129(Pt 7), 1822–1832.
PubMed Abstract | Google Scholar
Leh, S. E., Mullen, K. T., and Ptito, A. (2006b). Absence of S-cone input in human blindsight following hemispherectomy. Eur. J. Neurosci. 24, 2954–2960.
PubMed Abstract | Google Scholar
Marzi, C., Tassinari, G., Aglioti, S., and Lutzemberger, L. (1986). Spatial summation across the vertical meridian in hemianopics: a test of blindsight. Neuropsychologia 24, 749–758. doi: 10.1016/0028-3932(86)90074-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Miller, J. (1982). Divided attention: evidence for coactivation with redundant signals. Cogn. Psychol. 14, 247–279. doi: 10.1016/0010-0285(82)90010-X
CrossRef Full Text | Google Scholar
Miniussi, C., Girelli, M., and Marzi, C. A. (1998). Neural site of the redundant target effect electrophysiological evidence. J. Cogn. Neurosci. 10, 216–230. doi: 10.1162/089892998562663
CrossRef Full Text | Google Scholar
Pare, M., and Wurtz, R. (1997). Monkey posterior parietal cortex neurons antidromically activated from superior colliculus. J. Neurophysiol. 78, 3493–3497.
PubMed Abstract | Google Scholar
Poliva, O., Bestelmeyer, P. E., Hall, M., Bultitude, J. H., Koller, K., and Rafal, R. D. (2015). Functional mapping of the human auditory cortex: fMRI investigation of a patient with auditory agnosia from trauma to the inferior colliculus. Cogn. Behav. Neurol. 28, 160-180. doi: 10.1097/WNN.0000000000000072
PubMed Abstract | Google Scholar | Volledige Tekst | Google Scholar
Posner, M., Snyder, C., and Davidson, B. (1980). Aandacht en de detectie van signalen. J. Exp. Psychol. Gen. 109, 160-174. doi: 10.1037/0096-3445.109.2.160
CrossRef Full Text | Google Scholar
Raab, D. H. (1962). Statistical facilitation of simple reaction times. Trans. N. Y. Acad. Sci. 24, 574-590. doi: 10.1111/j.2164-0947.1962.tb01433.x
CrossRef Full Text | Google Scholar
Reuter-Lorenz, P., Nozawa, G., Gazzaniga, M. S., and Hughes, H. C. (1995). Fate of neglected targets: a chronometric analysis of redundant target effects in the bisected brain. J. Exp. Psychol. Hum. Percept. Perform. 21, 211-230. doi: 10.1037/0096-1523.21.2.211
PubMed Abstract | CrossRef Full Text | Google Scholar
Savazzi, S., and Marzi, C. A. (2004). The superior colliculus subserves interhemisferic neural summation in both normals and patients with a total section or agenesis of the corpus callosum. Neuropsychologia 42, 1608-1618. doi: 10.1016/j.neuropsychologia.2004.04.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Schiller, P. H., Malpeli, J. G., and Schein, S. J. (1979). Composition of geniculostriate input ot superior colliculus of the rhesus monkey. J. Neurophysiol. 42, 1124-1133.
PubMed Abstract | Google Scholar
Sommer, M., and Wurtz, R. (2000). Samenstelling en topografische organisatie van signalen uitgezonden van het frontale oogveld naar de superieure colliculus. J. Neurophysiol. 83, 1979–2001.
PubMed Abstract | Google Scholar
Song, J. H., Rafal, R. D., and McPeek, R. M. (2011). PNAS Plus: deficits in reach target selection during inactivation of the midbrain superior colliculus. Proc. Natl. Acad. Sci. U.S.A. 108, E1433–E1440. doi: 10.1073/pnas.1109656108
PubMed Abstract | CrossRef Full Text
Tomaiuolo, F. (1997). Blindsight in hemispherectomized patients as revealed by spatial summation across the vertical meridian. Brain 120, 795–803. doi: 10.1093/brain/120.5.795
PubMed Abstract | CrossRef Full Text | Google Scholar
Zhaoping, L. (2016). From the optic tectum to the primary visual cortex: migration through evolution of the saliency map for exogenous attentional guidance. Curr. Opin. Neurobiol. 40, 94–102. doi: 10.1016/j.conb.2016.06.017
PubMed Abstract | CrossRef Full Text | Google Scholar