Er zijn stambomen en populaties waar het principe van panmictische paring in wezen wordt doorbroken. Meestal gaat het om geografisch of sociaal betrekkelijk geïsoleerde kleine gemeenschappen. In deze gemeenschappen kan inteelt, d.w.z. consanguine paring van verschillende orden (een ouder – een kind, broers en zussen, neven en nichten, oom – nicht, enz.
Het probleem van de invloed van inteelt op de bevolking is het belangrijke probleem van de genetica 1. Nu heeft dit probleem nog geen definitieve oplossing gekregen.
In de meeste gevallen houdt de inteelt in een stamboom rekening met drie generaties. Het zijn de meest verre verwanten waarmee rekening wordt gehouden bij consanguine paringen is er sprake van achterneven en achternichten.
De inteelt wordt in aanmerking genomen met behulp van de inteeltfactor.
De inteeltfactor is de kans dat twee allelen van oorsprong identiek zijn.
Laten we eens kijken naar de methode van berekening van de inteeltfactor in een stamboom aangeboden door Wright 1, 2.
Voor het vinden van de inteeltfactor is het nodig:
om voor de huwende nakomelingen alle wegen in een stamboom te vinden die via de gemeenschappelijke voorouder deze nakomelingen verbinden.
om op elke weg het aantal stappen (afwisseling van generaties) op te tellen.
Dan is de inteeltfactor gelijk:
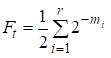 ……(1)
……(1)
waarbij r het aantal wegen is, m1, m2, … mi-aantal stappen op elke manier.
Wederom zullen we opmerken dat het aantal toevoegingen in de som (1) gelijk is aan het aantal manieren die via de gemeenschappelijke voorouders huwelijken met elkaar verbinden.
Laten we eens kijken naar concrete gevallen van stambomen, figuur 1. In deze figuur betekent het vierkant een mannelijk individu de cirkel – vrouwelijk individu. Stippellijnen en identieke cijfers geven mogelijke manieren in een stamboom die door de gemeenschappelijke voorouder (de man of de vrouw) verbinden van de nakomelingen consanguine trouwen. Op sommige plaatsen van wegen, Figuur 1с, zijn er twee cijfers 1 en 2. Het geeft aan dat dergelijke sites de gemeenschappelijke zijn voor de verschillende manieren.
Het huwelijk een vader – dochter is incest, Figuur 1а. De nakomeling (vrouw) is verbonden met de voorouder (man) daar 1 weg met één stap, d.w.z. m = 1. Er is dus een inteeltfactor.
Ft = 1/2.2-1 = 1/4
Er is een huwelijk broer – zus (sibs), Figuur 1b. Cijfers geven het aantal van de weg aan. Het aantal identieke cijfers op de weg van de huwende man naar de vrouw is gelijk aan het aantal stappen.
Laat elke nakomeling die gehuwd is zich op twee manieren verbinden met partner met twee stappen op de elke weg, dus m = 2. Vandaar dat er inteeltfactor is.
Ft = 1/2.(2-2 + 2-2) = 1/4
Het huwelijk tussen neven en nichten is weergegeven op figuur 1с.
Figuur 1. Enkele vormen van inteelt in een stamboom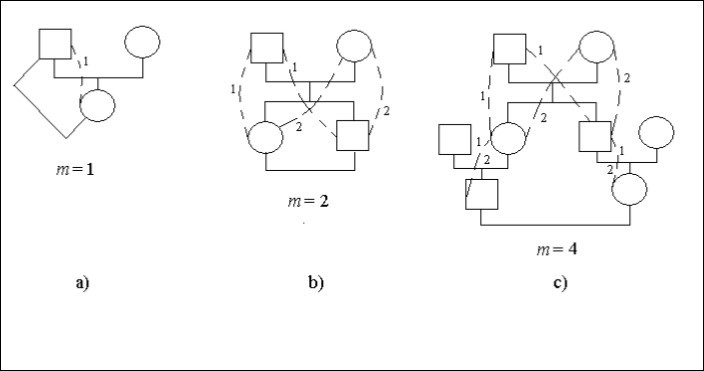
Elke nakomeling die getrouwd is, verbindt zich met partner de 2 manieren met 4 stappen op de elke manier, d.w.z. m = 4. Er is dus sprake van een inteeltfactor.
Ft = 1/2.(2-4 + 2-4) = 1/16
De formule (1) is gemakkelijk te begrijpen uit de volgende logica.
Het is duidelijk dat er 1/2 gen van het kind gemeenschappelijk is met elke ouder, 1/4 – met de grootvader en grootmoeder, 1/8 – met de overgrootvader en overgrootmoeder, enz. Als van de vrouw naar een voorouder de weg leidt met η stappen het aandeel van de gemeenschappelijke genen met deze voorouder is gelijk 2-η. Als van de man naar een willekeurige voorouder de weg leidt met stappen van μ is het aandeel van de gemeenschappelijke genen gelijk aan 2-μ. Bij het huwelijk van de man en de vrouw met de gemeenschappelijke voorouder zal het aandeel van de gemeenschappelijke genen met deze voorouder gelijk zijn aan 2-η. 2-μ = 2-m waarbij m = η + μ is het totaal aantal stappen door de gemeenschappelijke voorouder van de vrouw tot aan het huwelijk van de man. De grootte 1/2 2-m geeft de kans dat het gen dat toevallig bij de vrouw wordt gekozen van oorsprong identiek zal zijn aan het gen dat toevallig bij de man wordt gekozen. Dezelfde maat geeft de bijdrage aan de inteeltfactor Ft één weg door de gemeenschappelijke voorouder van de vrouw tot de man die trouwt.
Bijv. als de gemeenschappelijke voorouder op η = μ = 5 generaties verwijderd zal zijn van het huwelijk, is de kans dat zij het gemeenschappelijke gen hebben gelijk aan. 1/2 2-(5+5) = 1/2048 Deze grootte bij de analyse van de paring kan worden verwaarloosd. (Tabel 1)
Tabel 1.inteeltfactoren voor de enkele consanguine huwelijken zijn daarnaast weergegeven.
| De aard van het consanguine huwelijk | Inteeltfactor Ft |
| 1/32 | |
| Nichtoom – niece | 1/32 |
| Third cousins | 1/64 |
The inbreeding factor Ft is the probability of two alleles in posterity are identical by origin i.e. occur from same gene of ancestor 1. Or, that too most, the inbreeding factor is a probability of the allele origin in posterity is consanguineous. Hence, the probability of the non-consanguineous origins of identical alleles in homologous chromosomes is equal 1 – Ft.
Let’s find total probability of the recessive homozygote aa occurrence (eventаа) at random mating for an autosomal genes 3:
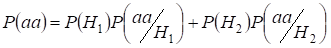 …….(2)
…….(2)
This formula follows from the following logic of mating: de oorsprong van identieke allelen in homologe autosomen is niet-consanguine (de kans op deze hypothese is P(H1) = 1- Ft )ontstaat ook homozygoteаа (voorwaardelijke kans op deze gebeurtenis is P(aa/H1) = q2) of oorsprong van identieke allelen is consanguine (de kans op deze hypothese is P(H2) = Ft ) en ontstaat homozygoteаа (voorwaardelijke kans op deze gebeurtenis is P(aa/H2) = q). De waarschijnlijkheid q van het optreden van de homozygoot aa in het laatste geval wordt bepaald door het feit dat als allel a in één autosoom voorkomt, bij consanguine oorsprong van de homozygoot hetzelfde allel noodzakelijkerwijs ook in het homologe autosoom moet voorkomen.
Hieruit volgt dat de totale kans dat de recessieve homozygoot voorkomt gelijk is aan:
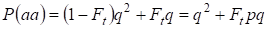 ……….(3)
……….(3)
waarbij in aanmerking wordt genomen dat p = 1 – q er de dominante allel a frequentie is.
Zo is er ook de totale kans dat de dominante homozygoot AA voorkomt bij een toevallige paring:
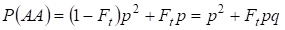 ……….. (4)
……….. (4)
Dus, volgens (3) en (4) neemt de totale frequentie van homozygoten (dominant en recessief) in de ingeteelde stamboom toe met 2Ftpq. Vandaar dat de frequentie van heterozygoten afneemt bij gelijke grootte en gelijk wordt aan 2pq – 2pqFt = 2pq(1-Ft) omdat de som van alle zygote frequenties gelijk is aan 1.
Op grond van de inteeltfactor en ook (3) en (4) moet de Hardy – Weinberg wet in de volgende generatie van inteelt stamboom voor een autosomaal genoom worden opgeschreven de verdelingen van zygote genotypen als 4:
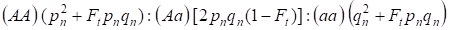 …………(5)
…………(5)
Waarbij pn de dominante allelen a-frequentie in generatie n is, qn – de recessieve allelen a-frequentie, dus pn + qn = 1.
De ratio (5) verwijst ook naar Wright’s ratio 5. Wright’s ratio toont aan dat in een ingeteelde stamboom na de consanguine paring van individuen de volgende effecten worden waargenomen: er is toename van de autosomale homozygoten fractie in een stamboom op Ft pnqnen afname van de autosomale heterozygoten fractie in (1-F1) in vergelijking met een panmictische populatie.
Maar al na één generatie is het Hardy – Weinberg evenwicht hersteld dat het geringe belang van de afzonderlijke consanguine paring voor een stamboom specificeert:
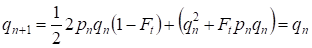 ………(6)
………(6)
Het is duidelijk dat voor een populatie als geheel de afzonderlijke daad van de consanguine paring nog geringer van belang zal zijn dan voor een stamboom.
Hetzelfde effect van variatie in de frequentie van zygote genotypen wordt waargenomen en voor geslachtsgebonden genen.
De verdeling van de genotypen gekoppeld aan het Х-chromosoom in een panmictische stamboom wordt door de wet van Hardy – Weinberg beschreven als 1:
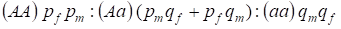 ………..(7)
………..(7)
De frequentie van een dominant allel A zullen we aanduiden bij de mannen pm en bij de vrouwen pf. Voor recessieve allelenesa is dat qm en qf.
Bij de paring is er een verhouding van genotypen bij de vrouwen volgens het product (pf + qf) (pm +qm).
Laat ons de totale waarschijnlijkheid vinden van het voorkomen van de recessieve homozygoot aa bij een willekeurige paring voor de genen van de dochter die gekoppeld zijn aan hetХ-chromosoom volgens de formule (2). De logica van de gebeurtenissen is vergelijkbaar met die voor autosomen, behalve dat de voorwaardelijke kans dat een genotypeаа voorkomt uit de niet-consanguine allelen volgens (7) gelijk is aan P(aa/H1) = qmnqfn. Voor de consanguine allelen is de voorwaardelijke kans op het voorkomen van een genotypeаа gelijk aan P(aa/H2) = qf(n+1). Bij dochters, d.w.z. in generatie n+1 wordt de kans op het voorkomen van een homozygoot aa bepaald door het feit dat als allel a zich in één Х-chromosoom bevindt, bij consanguine oorsprong van de homozygoot hetzelfde allel noodzakelijkerwijs ook in het homologe Х-chromosoom aanwezig moet zijn.
Hieruit volgt dat de totale kans dat de recessieve homozygoot voorkomt gelijk is:
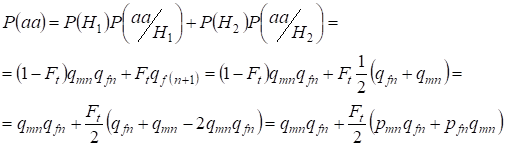 ……(8)
……(8)
waarbij pmn de dominante allelen A-frequentie in generatie n bij mannen is en pfn – bij vrouwen. Voor de recessieve allelen a dienovereenkomstig qmn en qfn . Daarnaast wordt qmn =1 – qmn en pfn = 1 – qfn.
Ook voor het vinden van de totale kans op het voorkomen van de dominante homozygoot wordt gebruik gemaakt van:
 ,
,
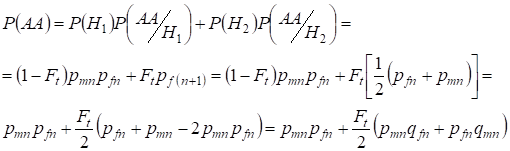 ……….(9)
……….(9)
en een heterozygoot P(Aa) = 1- P(aa) – P(AA) vinden we de verdeling van de genotypen in Х-chromosomen van de ingeteelde stamboom bij de paring:
 ……..(10)
……..(10)
Het is tegelijkertijd aangetoond dat zowel in de autosomen als in de inteelt de additieven in dominante en recessieve homozygote frequenties in de geslachtsgebonden chromosomen identiek zijn.
Een andere manier om de verhouding (10) te vinden zal bestaan uit het volgende.
Laten we eerst de totale kans vinden dat een heterozygoot Aa voorkomt, d.w.z. P(Aa) bij de gelegenheid van de willekeurige consanguine paring. Deze kans volgt uit de volgende logica van de paring: de oorsprong van de allelen is niet-consanguine (de kans op deze hypothese is P(H1) = 1 – Ft ) ontstaat ook heterozygoot Aа (de voorwaardelijke kans op deze gebeurtenis is P(Aa/H1) = pmnqfn + pfnqmn, zie formule (7)) of de oorsprong van de allelen is consanguine (de waarschijnlijkheid van deze hypothese is P(H2) = Ft) en ontstaat de heterozygoot Aа (voorwaardelijke waarschijnlijkheid van deze gebeurtenis is gelijk aan nul P(Aa/H2) = 0 ). De laatste uitspraak wordt bepaald door het feit dat bij consanguine oorsprong gelijksoortige allelen in homologe Х-chromosomen niet kunnen ontstaan uit heterozygoteАа bestaande uit verschillende allelen.
Dus, met behulp van de formule van de totale waarschijnlijkheid vinden we de totale waarschijnlijkheid van het optreden van de heterozygote Aa:
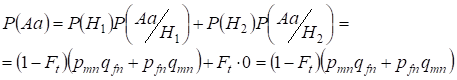 …….(11)
…….(11)
De frequentie van heterozygoten Aa neemt af, zoals eerder is aangetoond, door een identieke toename van de dominante AA- en recessieve aa-homozygotenfrequentie. Zo is de toename van de recessieve aa-homozygotenfrequentie gelijk:
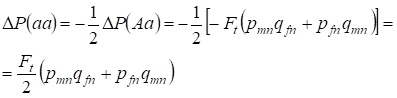 ……..(12)
……..(12)
waarbij afname van de heterozygoten-Aa-frequentie ten opzichte van de panmictische stamboom volgens (7) en (11) gelijk is. ΔP(Aa) = – Ft (pmnqfn + pfnqmn) Met behulp van (11) en (12) om (7) bij te werken voor gevallen van inteelt vinden we de verdeling van genotypen (10) in Х-chromosomen in de dochters inteeltstamboom.
In de volgende generatie is de frequentie van de recessieve allelen a bij vrouwen gelijk:
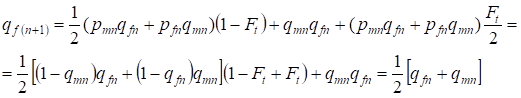 ……..(13)
……..(13)
Dat komt volledig overeen met dezelfde frequentie bij afwezigheid van inteelt, d.w.z. er is herstel van het Hardy – Weinberg evenwicht of is nauwkeuriger benadering van het evenwicht.
De verhoudingen (7) en (13) laten als geheel zien dat de allelfrequenties in een stamboom voor de rekening van inteelt niet variëren. Er is alleen een herverdeling van allelen. Een deel van de allelen verlaat de heterozygoten en gaat over naar de homozygoten.