1. Cilium
2. flagellum
3. struktura
4. ruch
5. Formacja
Mikrotubule są składnikami cytoszkieletu o ważnych funkcjach w fizjologii komórki. Rusztowanie mikrotubul w cytoplazmie jest wysoce plastyczne dzięki zdolności mikrotubul do polimeryzacji i depolimeryzacji. Jednak nie wszystkie mikrotubule komórkowe podlegają tym stadiom kurczenia się lub wzrostu. Cilia, flagella i centriole/ciała podstawowe to struktury komórkowe zawierające bardzo stabilne (liczba i długość) i wysoko zorganizowane mikrotubule. Na tej stronie zajmiemy się rzęskami i flagellami.
1. Cilium
Cilium to cienkie i długie wypustki komórkowe o średnicy ok. 0,25 µm i długości ok. 10-15 µm, występujące w komórkach zwierzęcych i niektórych jednokomórkowych gatunkach eukariotycznych. Zazwyczaj są one ciasno upakowane na wolnej powierzchni komórek nabłonkowych (ryc. 1 i 2), takich jak nabłonek dróg oddechowych, nabłonek przewodów rozrodczych, skrzela ryb i małży itp. Cilia jest strukturą ruchliwą, a jej główną funkcją jest przemieszczanie otaczającego ją płynu, np. śluzu na powierzchni dróg oddechowych, wody wokół nabłonka skrzeli, ale także oocytu w przewodzie jajowodowym samicy. Wiele organizmów jednokomórkowych może poruszać się napędzanych przez rzęski, a inne mogą ich używać do generowania wirów wodnych w celu łapania pokarmu. Rzęski węzłowe embrionu są zaangażowane w inicjowanie osi lewo-prawo podczas rozwoju embrionalnego kręgowców. Ruch rzęsek jest jak bicie, które pobudza ciecz równolegle do powierzchni komórki.
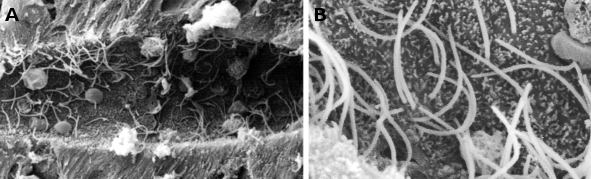
Rysunek 1. Obrazy skaningowej mikroskopii elektronowej przedstawiające kanał centralny rdzenia kręgowego minoga. Można zaobserwować wiele rzęsek (w większym powiększeniu w B) i małe mikrowypustki w apikalnej domenie komórek ependimalnych.
Obraz 2. Obrazy nabłonka oddechowego w transmisyjnej mikroskopii elektronowej. Komórki z wyraźną cytoplazmą wykazują wiele rzęsek na ich powierzchni wierzchołkowej.
Istnieją rzęski, które nie mogą się poruszać, a zatem nie są przeznaczone do przemieszczania się w płynie. Te rzęski są znane jako rzęski pierwotne. Większość badanych dotychczas komórek (z wyjątkiem krwinek czerwonych) posiada rzęski pierwotne: komórki jajowodów, neurony, chondrocyty, komórki ektodermy, komórki mezenchymalne, komórki nabłonka układu moczowego, hepatocyty, a nawet komórki hodowlane. Początkowo uważano, że rzęski pierwotne to rzęski niefunkcjonalne. Jednak w błonie rzęskowej znaleziono wiele typów receptorów i kanałów jonowych, dlatego uznano je za struktury czuciowe komórki. Na przykład, receptory węchowe znajdują się w rzęskach ich dendrytów, a zewnętrzne segmenty pręcików i czopków siatkówki są właściwie zmodyfikowanymi rzęskami. Niektóre receptory s± bardziej upakowane w błonie rzęskowej niż w innych domenach błony plazmatycznej. Ponadto, we wnętrzu rzęsek znajduje się wiele różnych cząsteczek biorących udział w transdukcji sygnału. Wyższy stosunek powierzchni do objętości rzęski sprawia, że wewnątrzrzęskowe odpowiedzi molekularne s± bardziej intensywne i wydajne niż gdyby znajdowały się na zewn±trz rzęski. Oprócz sygnału chemicznego, rzęski pierwotne mogą wykrywać ruch płynu na zewnątrz komórki i działać jako mechanoreceptory.
2. Flagellum
Flagella są podobne do rzęsek, ale są znacznie dłuższe, ok. 150 µm, i nieco grubsze. W komórkach występują znacznie mniej licznie niż rzęski. Główną funkcją flagelli jest poruszanie komórką. Ruch flagellum różni się od ruchu cilium tym, że kierunek ruchu jest prostopadły do powierzchni komórki (a nie równoległy), czyli zgodny z kierunkiem osi podłużnej flagellum. Flagellę można często zaobserwować w komórkach ruchliwych, takich jak organizmy jednokomórkowe i plemniki.
Struktura
Cilia i flagella są złożonymi strukturami zawierającymi ponad 250 różnych białek. Obie mają tę samą centralną organizację mikrotubul i inne związane z nimi białka, łącznie znane jako akson, i ograniczone przez błonę plazmatyczną (Rysunek 3). Poza aksonemem, wewnątrz rzęsek/flagelli znajduje się wiele rozpuszczalnych cząsteczek tworzących macierz. Akson składa się z 9 par mikrotubul otaczających inną, centralną parę mikrotubul. Organizację tę można zapisać jako (9 x 2) + 2. W rzęskach pierwotnych brak jest centralnej pary mikrotubul. Każda mikrotubula pary centralnej zbudowana jest z 13 protofilamentów, ale mikrotubule par peryferyjnych dzielą między sobą niektóre protofilamenty. Tak więc para peryferyjna jest utworzona przez mikrotubule A i B. Mikrotubula A zawiera 13 protofilamentów, a mikrotubula B zawiera 10 lub 11 protofilamentów, dzieląc 2 lub 3 z mikrotubulą A.

Rysunek 3. Główne składniki molekularne rzęsek i flagelli. W rzęskach pierwotnych para centralna jest nieobecna.
Organizacja mikrotubul w aksonemie jest wynikiem działania rusztowania białek. Znaleziono już dwanaście białek jako składniki aksonu zaangażowane w utrzymanie organizacji mikrotubul. Sąsiednie pary mikrotubul obwodowych są połączone między sobą za pomocą nexin. W każdej parze, mikrotubula A jest poł±czona szprychami białkowymi z centralnym pierścieniem, który zawiera centraln± parę lub mikrotubule. Dineina jest białkiem motorycznym związanym z peryferyjnymi mikrotubulami zaangażowanymi w ruch rzęsek i flagelli.

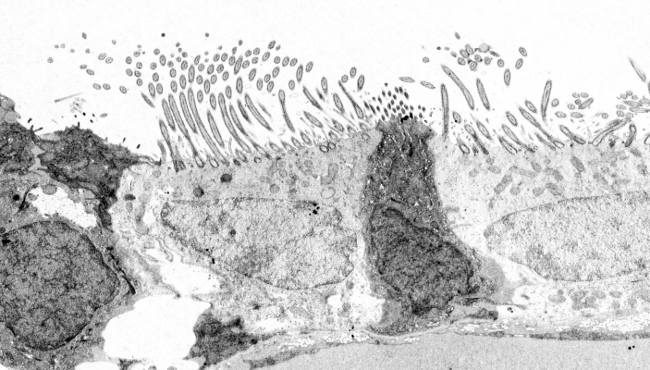
Figura 4. Ultrastruktura cilium komórki ependimalnej rdzenia kręgowego. (9+2)x2 oznacza 9 par obwodowych i 1 centralną parę mikrotubul.
Mikrotubule są polimeryzowane z ciałek podstawnych (ryc. 3 i 4). Ciało bazalne zbudowane jest z 9 potrójnych mikrotubul tworzących cylinder (podobnie jak centriole). W każdym triplecie tylko jedna mikrotubula (mikrotubula A) posiada kompletny zestaw protofilamentów, natomiast mikrotubule B i C dziel± czę¶ć z nich między siebie. Od ciała podstawowego mikrotubule A i B rozrastaj± się i tworz± mikrotubule peryferyjne aksonu. Tuż nad ciałem podstawowym znajduje się obszar rzęsek zwany strefą przejściową, w której znajduje się 9 par obwodowych i brak pary centralnej. Bezpośrednio za strefą przejściową znajduje się blaszka podstawna, z której polimeryzuje centralna para mikrotubul, tworząc aksonem. Wszystkie mikrotubule mają koniec plusowy skierowany w stronę wierzchołka cilium/flagellum. Proksymalny koniec ciała podstawowego (wewnętrzny, czyli minusowy koniec mikrotubul) jest zakotwiczony do cytoszkieletu komórki przez długie włókna białkowe zwane korzonkami rzęskowymi
Poza aksonemem rzęski/flagellum mają inne przedziały. Błona zawiera wiele receptorów i kanałów umożliwiających wyczuwanie środowiska, szczególnie w rzęskach pierwotnych. Faza płynna wnętrza nazywana jest macierzą, która pomaga w utrzymaniu organizacji całej struktury i jest odpowiedzialna za przekazywanie informacji zebranych przez receptory błonowe. Innymi wyróżniającymi się obszarami są ciało bazalne znajdujące się u podstawy oraz część wierzchołkowa rzęski/flagellum, która zawiera białka stabilizujące plusowe końce mikrobuli.
4. Ruch
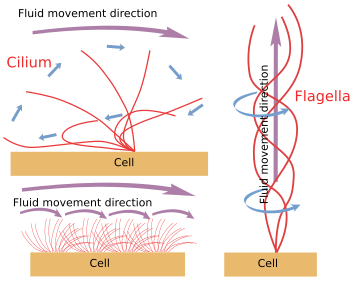
Rysunek 6. Modele ruchu cilium i flageli. Generują one różne kierunki ruchu płynu.
Gdy cilium/flagella są mechanicznie oderwane od komórki, poruszają się aż do wyczerpania zapasów ATP. Oznacza to, że mechanizm ruchu jest samoistny (rysunek 6). W rzeczywistości ruch jest wytwarzany przez nasuwanie się peryferyjnej pary mikrotubul na sąsiednią. Nexin i białka spoke zapobiegają dezorganizacji aksonu, ale umożliwiają te ruchy. Dineina jest białkiem motorycznym odpowiedzialnym za ruch ślizgowy. Energia dostarczana jest przez ATP. Dineiny s± zakotwiczone czę¶ci± globularn± do mikrotubuli A jednej pary obwodowej, a czę¶ci± ogonow± do mikrotubuli B pary przylegaj±cej. Mechanizm molekularny jest podobny jak w przypadku dinein cytozolowych, ale zamiast transportować ładunek, poruszaj± one mikrotubule. Dla efektywnego ruchu niezbędna jest koordynacja działania dinein aksonu. Fale wapniowe we wnętrzu rzęsek/flagelli mog± koordynować aktywację dinein i w razie potrzeby zmieniać częstotliwo¶ć ruchu. Należy zauważyć, że nie wszystkie dineiny muszą być aktywowane w tym samym czasie, ale synchronicznie.
5. Formacja
Podczas różnicowania, komórki wytwarzają wszystkie niezbędne dla ich prawidłowej fizjologii rzęski i chorągiewki. Oznacza to, że wszystkie one muszą być wytworzone od podstaw. Mikrotubule aksonów są zarodkowane z mikrotubul A i B ciałek podstawnych, więc na jedną rzęskę/flagellę przypada jedno ciałko podstawne. Jak powstają liczne ciałka podstawne? Istnieją co najmniej trzy sposoby wytwarzania ciałek podstawnych: a) przy użyciu centrioli jako szablonów do zarodkowania ciałek podstawnych; b) z bezpostaciowego materiału znanego jako deuterosom; c) u roślin istnieją wyraźne agregaty białkowe, które mogą zarodkować ciałka podstawne.
Liczne ludzkie patologie są konsekwencją wad cilium/flagelli. Są one znane jako ciliopatie i obejmują przypadkową lateralizację, nieprawidłowe zamknięcie cewy nerwowej, polidaktylię, torbielowatość nerek, patologie wątroby i trzustki, degenerację siatkówki, otyłość i defekty poznawcze.
Bibliografia
Marshall WF, Nonaka S. 2006. Cilia: tuning in to the cell’s antenna. Current biology. 16:R604-R614.
Satir P, Christensen ST. 2007. Overview of structure and function of mammalian cilia. Annual review of phisiology. 69:377-400.