1. Cilium
2. Flagellum
3. Struktur
4. Rörelse
5. Bildning
Mikrotubuli är cytoskelettkomponenter med viktiga funktioner i cellfysiologin. Cytoplasmens mikrotubulära ställning är mycket plastisk tack vare mikrotubulernas polymerisations- och depolymeriseringskapacitet. Det är dock inte alla cellmikrotubuli som befinner sig under detta krympnings- eller tillväxtstadium. Cilior, flageller och centrioler/basala kroppar är cellulära strukturer som innehåller mycket stabila (antal och längd) och mycket organiserade mikrotubuli. På den här sidan behandlar vi cilier och flageller.
1. Cilium
Cilier är tunna och långa cellutsprång med en diameter på cirka 0,25 µm och en längd på cirka 10 till 15 µm, som finns i djurceller och vissa encelliga eukaryota arter. De är vanligtvis tätt packade på den fria ytan av epitelceller (figurerna 1 och 2), t.ex. epitelet i luftvägarna, epitelet i fortplantningsgångarna, gälarna hos fiskar och tvåskaliga djur och så vidare. Cili är rörliga strukturer och deras huvudsakliga funktion är att förflytta den omgivande vätskan, t.ex. slem på luftvägarnas yta, vatten runt gälarnas epitel, men också oocyten i kvinnans äggledare. Många encelliga organismer kan förflytta sig med hjälp av cilier, och andra kan använda dem för att skapa vattenvirvlar för att fånga föda. Embryonala nodala cilier har involverats i initieringen av vänster-höger-axeln under embryonalutvecklingen hos ryggradsdjur. Ciliernas rörelse är som ett slag, som impulserar vätskan parallellt med cellytan.
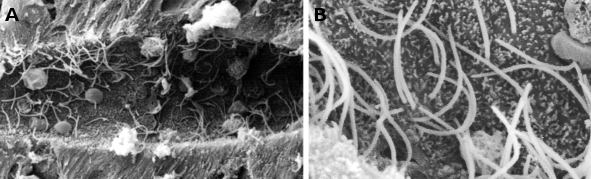
Figur 1. Skanningselektronmikroskopiska bilder som visar den centrala kanalen i en nejonögons ryggmärg. Många cilier kan observeras (i högre förstoring i B) och små mikrovilli vid den apikala domänen av ependimala celler.
Figur 2. Transmissionselektronmikroskopiska bilder av det respiratoriska epitelet. Celler med klar cytoplasma visar många cilier i sin apikala yta.
Det finns cilier som inte kan röra sig och därför är de inte avsedda för vätskeförflyttning. Dessa cilier är kända som primära cilier. De flesta celler som studerats hittills (utom röda blodkroppar) bär primära cilier: äggledarceller, neuroner, kondrocyter, ektodermceller, mesenkymala celler, urinepitelceller, hepatocyter och till och med odlade celler. Ursprungligen betraktades primära cilier som icke-funktionella cilier. Många receptortyper och jonkanaler fanns dock i ciliarmembranet, så de betraktades som cellsensoriska strukturer. Till exempel finns luktreceptorer i dendriternas cilier, och de yttre segmenten av stavar och kottar i näthinnan är faktiskt modifierade cilier. Vissa receptorer är mer packade i ciliarmembranet än i andra plasmamembranområden. Dessutom finns det en mängd olika molekyler i ciliernas inre som är involverade i signalöverföringsroller. Det högre förhållandet mellan yta och volym i ett cilium gör de intraciliära molekylära reaktionerna mer intensiva och effektiva än om de fanns utanför ciliet. Förutom kemiska signaler kan primära cilier upptäcka vätskeförflyttningar utanför cellen och fungera som mekanoreceptorer.
2. Flagellum
Flagella liknar cilier, men de är mycket längre, cirka 150 µm långa, och något tjockare. De är betydligt mindre talrika än cilier i cellerna. Flagellas huvudsakliga funktion är att förflytta cellen. Flagellans rörelse skiljer sig från ciliums eftersom rörelseriktningen är vinkelrät mot cellytan (inte parallell), det vill säga riktningen för flagellans längdaxel. Flageller kan ofta observeras i rörliga celler som encelliga organismer och spermier.
Struktur
Cilier och flageller är komplexa strukturer som innehåller mer än 250 olika proteiner. Båda delar samma centrala mikrotubuliorganisation och andra associerade proteiner, som tillsammans kallas axonem, och begränsas av plasmamembranet (figur 3). Förutom axonem finns det många lösliga molekyler inuti cilierna/flagellorna som utgör matrisen. Axoneme består av nio par mikrotubuli runt ett annat centralt par mikrotubuli. Denna organisation kan skrivas som (9 x 2) + 2. Primära cilier saknar ett centralt par mikrotubuli. Varje mikrotubuli i det centrala paret består av 13 protofilament, men mikrotubuli i de perifera paren delar några protofilament med varandra. Ett perifert par bildas alltså av A- och B-mikrotubuli. A-mikrotubuli innehåller 13 protofilamenter och B-mikrotubuli innehåller 10 eller 11 protofilamenter som delar 2 eller 3 med A-mikrotubuli.

Figur 3. Huvudsakliga molekylära komponenter i cilier och flageller. I primära cilier saknas det centrala paret.
Axonemens mikrotubuliorganisation är resultatet av en ställning av proteiner. Tolv proteiner har redan hittats som beståndsdelar i axonemet som är involverade i upprätthållandet av mikrotubuliorganisationen. De angränsande perifera mikrotubuliparen är förbundna mellan varandra genom nexin. I varje par är A-mikrotubulerna förbundna med proteinekar till en central ring som innehåller det centrala paret eller mikrotubulerna. Dinein är ett motorprotein som är knutet till de perifera mikrotubuli som är involverade i rörelsen av cilier och flageller.

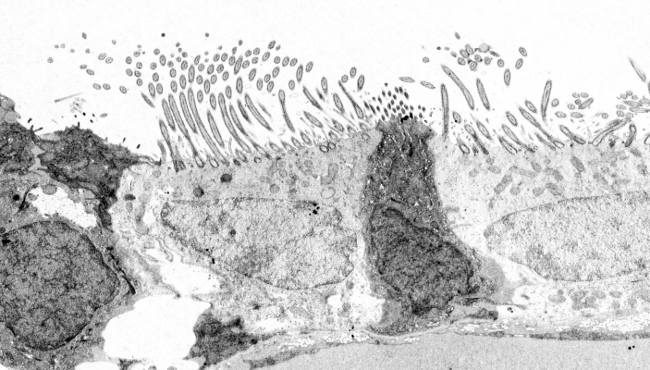
Figur 4. Ultrastruktur av ett cilium i en ependimal cell i ryggmärgen. (9+2)x2 betyder 9 perifera par och 1 centralt par mikrotubuli.
Mikrotubuli polymeriseras från basala kroppar (figurerna 3 och 4). Basalkroppen består av 9 trippelmikrotubuli som bildar en cylinder (liknar centrioler). De saknar ett centralt par mikrotubuli, så det är (9×3)+0. I varje triplett har endast en mikrotubuli (A-mikrotubuli) en komplett uppsättning protofilament, medan B- och C-mikrotubuli delar några av dem mellan varandra. Från basalkroppen växer A- och B-mikrotubuli och bildar axonemets perifera mikrotubuli. Strax ovanför basalkroppen finns ett område av cilierna som kallas övergångszon och som innehåller de nio perifera paren och inget centralt par. Omedelbart efter övergångszonen finns den basala plattan, från vilken det centrala paret mikrotubuli polymeriseras för att fullborda axonemet. Alla mikrotubuli har plusändan mot cilium/flagellums spets. Den proximala änden av basalkroppen (den inre, eller mikrotubulernas minusände) är förankrad i cellens cytoskelett genom långa proteinfibrer som kallas ciliary rootlets
När axoneme har cilier/flageller andra avdelningar. Membranet innehåller många receptorer och kanaler för att känna av omgivningen, särskilt i primära cilier. Den flytande fasen i det inre kallas matris, som hjälper till att hålla hela strukturen organiserad och ansvarar för att överföra den information som samlas in av membranreceptorerna. Andra distinkta områden är den basala kroppen som ligger vid basen och den apikala delen av cilium/flagellum, som innehåller proteiner som stabiliserar mikrokulornas plusändar.
4. Rörelse
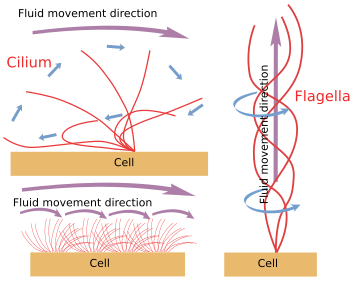
Figur 6. Modeller för cilium- och flagellarörelser. De genererar olika rörelseriktningar för vätskor.
När cilium/flagella lossas mekaniskt från cellen fortsätter de att röra sig tills ATP-lagret är uttömt. Det innebär att rörelsemekanismen är inneboende (figur 6). I själva verket produceras rörelsen genom att ett perifert par mikrotubuli glider över grannen. Nexin- och spokeproteiner förhindrar axonemets desorganisering, men möjliggör dessa rörelser. Dinein är det motorprotein som ansvarar för glidrörelsen. Energin tillhandahålls av ATP. Dinein förankras med sin globulära del till A-mikrotubuli i ett perifert par och med sin svansdel till B-mikrotubuli i det angränsande paret. Den molekylära mekanismen liknar den för de cytosoliska dineinerna, men i stället för att transportera en last flyttar den en mikrotubuli. För en effektiv förflyttning krävs en samordning av dinainerna i axonemet. Kalciumvågor i cilien/flagellans inre kan samordna aktiveringen av dineinerna och ändra rörelsefrekvensen vid behov. Det är viktigt att notera att alla dineiner inte måste aktiveras samtidigt, utan synkront.
5. Bildning
Vid differentiering producerar cellerna alla nödvändiga cilier och flageller för sin normala fysiologi. Det innebär att alla måste genereras från grunden. Axoneme-mikrotubuli kärnbildas från A- och B-mikrotubuli från basalkropparna, så det behövs en basalkropp per cilium/flagellum. Hur bildas flera basalkroppar? Det finns minst tre sätt att producera basalkroppar: a) genom att använda centrioler som mallar för kärnbildning av basalkroppar, b) från amorft material som kallas deuterosom, c) i växter finns det distinkta proteinaggregat som kan bilda kärnor till basalkroppar.
Enormt många mänskliga patologier är en följd av brister i cilium/flagella. De kallas ciliopatier och omfattar slumpmässig lateralitet, felaktig stängning av neuralröret, polydaktyli, cystiska njurar, lever- och bukspottkörtelpatologier, näthinnedegeneration, fetma och kognitiva defekter.
Bibliografi
Marshall WF, Nonaka S. 2006. Cilia: att ställa in sig på cellens antenn. Aktuell biologi. 16:R604-R614.
Satir P, Christensen ST. 2007. Översikt över struktur och funktion hos däggdjurs cilier. Annual review of phisiology. 69:377-400.